
American Journal of Microbiology and Immunology, Vol. 1, No. 3, November 2016 Publish Date: Jan. 21, 2017 Pages: 32-39
A Dumpy Review on Preservation of Methicillin-Resistant Staphylococcus aureus (MRSA) in the Laboratory
Abdulhadi Sale Kumurya*
Department of Medical Laboratory Science, Faculty of Allied Health Sciences, Bayero University, Kano, Nigeria
Abstract
Staphylococcus aureus has emerged as one of the most important human pathogens, and has over the past several decades, been a leading cause of hospital and community acquired infections. Methicillin-resistant strains of S. aureus (MRSA) were first recognized in the 1980’s as a major clinical and epidemiological problem. Hospitals are still facing this problem today. Preservation of strains in a microbiology laboratory is of great importance for quality control, teaching, and research. Freezing is a very common method of preservation and storage of microorganisms. Studies concentrate on the viability of the microorganisms after a certain storage period. Little attention is given to the influence of storage conditions on characteristics of the stored strains. This article describes methods, reagents, and equipment commonly utilized for the growth, maintenance, and characterization of S. aureus in the laboratory. It also explains the methods of identification of S. aureus as methicillin resistant strains, and generated a method for freezer stock of S. aureus for long-term storage. The Microbank Bacterial Preservation system (Pro-lab Diagnostics) is a well-known system for freezer storage of all kinds of microorganisms and is used in laboratories all over the world. Staphylococcus aureus is a Biosafety Level 2 (BSL-2) pathogen. Follow all appropriate guidelines and regulations for the use and handling of pathogenic microorganisms. No special precautions are necessary when working with MRSA.
Keywords
Staphylococcus aureus, MRSA, Growth, Strain Selection, Freezer Stock
Received: December 9, 2016
Accepted: December 20, 2016
Published online: January 21, 2017
@ 2016 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY license. http://creativecommons.org/licenses/by/4.0/
Contents
1. Introduction 2. Tactical Preparations 2.1. Fundamental Procedure 1: Growing S. aureus on Solid Media 2.2. Fundamental Procedure 2: Testing for Methicillin Resistance in S. aureus 2.3. Fundamental Procedure 3: Growing of S. aureus in Liquid Media 2.4. Fundamental Procedure 4: Preparing Frozen Stocks of S. aureus 3. How to Culture Staphylococcus aureus on Liquid and Solid Media 3.1. Brain Heart Infusion (BHI) Agar Plates 3.2. Brain Heart Infusion (BHI) Broth 3.3. Mueller-Hinton Agar Plates Supplemented with 2% NaCl and Oxacillin (MHA-Salt Plates) 3.4. Chemically Distinct Culture Media 3.5. Sterile DMSO 4. Conclusion
1. Introduction
Staphylococcus aureuswas originally isolated from a knee abscess in 1881 [40]. Ever since then, S. aureus has become the most leading nosocomial infection in the developed world (i.e. the MRSA epidemic). Even though the frequency of HA-MRSA among hospital-acquired infections is at present declining, new subsets of MRSA (USA300, USA400, ST239, etc.) have arisen in the community setting [8]. These community-acquired MRSA (CA-MRSA) isolates exhibit increased ability to cause disease in healthy persons (Weber, 2005). The primary clinical presentation of USA300 is skin and soft-tissue infections, but it has also been established to cause fatal necrotizing pneumonia [35].
MRSA as a whole is proficient of causing a wide variety of clinical presentations, including: endocarditis, myocarditis, osteomyelitis, pneumonia, surgical site infections, toxic shock syndrome, septicemia and scalded skin syndrome. The capability of S. aureus to cause such varied diseases is mainly attributed to the diversity of virulence factors encoded in the S. aureus genome and the innate resistance of S. aureus to nearly all facets of the human immune system. One such virulence factor produced by S. aureus is the discrete yellowish-orange pigment called staphyloxanthin (aureus means "golden" in Latin). Pigment production has been shown to assist in the resistance of S. aureus to oxidative damage [27, 33, 41]. Additional main virulence factors encoded by S. aureus include: coagulases (coa and vwb), toxins (e.g. hla, hld, hlg, set8, psmα), proteases (e.g. spl, sspC, aur), lipases (lip and geh), adhesins (e.g. cflA, cflB, fnbA, fnbB), and factors involved in immune resistance (e.g. ldh1, hmp, katA, spa, eap, chp, sak, and capsule) [10, 15, 16].
Away from these obvious virulence factors, S. aureus is also a swift growing organism that is able of utilizing a variety of carbon sources (interestingly, USA300 isolates grow faster in vitro than their HA-MRSA counterparts [49]. As stated previously, all S. aureus strains show an in vitro auxotrophy for arginine and proline, although for proline, this is only during growth on glycolytic carbon sources (e.g. glucose). This is caused by catabolite-mediated suppression of the proline biosynthetic genes via the catabolite response regulator, CcpA [31]. Therefore, careful deliberation of the growth media chosen for testing should be taken, given that S. aureus exhibits wide-ranging metabolic regulation.
S. aureus is a gram-positive coccus (~0.6μm in diameter) that colonizes the skin and/or nares of most humans. Given access to other tissues (via tissue disruption or an impaired immune system), S. aureus can cause a wide variety of serious clinical manifestations including: skin and soft tissue infections (SSTI's), pneumonia, osteomyelitis, septicemia and endocarditis. Fundamental procedure (a) explains the culturing of S. aureus on agar plates, fundamental procedure (b) explains the Oxacillin-salt agar method for determining methicillin-resistance of S. aureus strains, fundamental procedure (c) explains the culturing of S. aureus in liquid media, and fundamental procedure (a) explains the preparation of a frozen stock of S. aureus from a liquid culture. Further information concerning unique molecular characteristics of CA-MRSA isolates (Table 1) and selective antibiotic concentrations (Table 2) are provided.
Table 1. Distinctive information concerning molecular characteristics of CA-MRSA isolates.

Table 2. Selective antibiotic concentrations for use with S. aureus cultures.
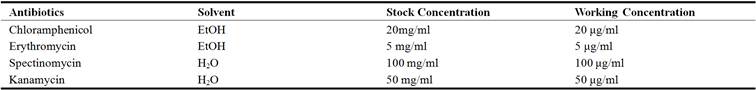
2. Tactical Preparations
Unlike other pathogenic bacteria studied in the laboratory, there are many generally utilized strains of S. aureus (Table 1). This is a proposal of the information that S. aureus strains show proof of distinction in commonly studied in vitro phenotypes, different degrees of virulence in mouse models of infection, and were isolated from a wide variety of infection sites and individuals. As stated earlier, S. aureus is normally divided into the methicillin-resistant (MRSA) and -susceptible subgroups (MSSA). MRSA strains were first isolated in the 1960's [26] and it was later on resolute their resistance was associated to the attainment of a mobile genetic element (staphylococcal cassette chromosome) harboring the mecA gene, (SCCmec) [50]. Ever since the recognition of the original SCCmec cassette, research on this subject has discovered the existence of other SCCmec cassette types that bestow extra antibiotic resistance genes to S. aureus isolates (for a description of the SCCmec types (IWG-SCC), 2009).
The universally studied MSSA isolates include: NCTC8325 (RN1), NCTC8325-4 (RN450), SH1000, RN6390, RN4220, and Newman. NCTC8325 (RN1) was isolated in 1960 from a sepsis patient and was mainly employed in the laboratory for the proliferation of phage 47 and the study of S. aureus genetics [38]. To create isolation of phage 47 easier, NCTC8325 was cured of three endogenous prophages determined in its genome via consecutive rounds of UV mutagenesis, yielding strain NCTC8325-4 (RN450) [19, 38]. Ultimately, it became obvious that NCTC8325 exhibited reduced toxin making, pigmentation, and virulence in mice, in contrast to other clinical S. aureus isolates. Genomic relationship of NCTC8325 to other MSSA strains confirmed that this strain had mutations in rsbU (activator of sigB) and tcaR (activator of spa) [19], making it a poor form for the study of S. aureus genetic regulation and virulence (albeit it has been generally utilized for the study of these two topics). To reverse this loss of virulence, the rsbU mutation was repaired in strain NCTC8325-4, yielding strain SH1000 [22]. Similarly, RN6390 is a more virulent derivative of NCTC8325-4 (produces greater amounts of exoprotein and α-toxin), but it was produced via sequential phage transduction [22].
Similar to SH1000 and RN6390, RN4220 is also a derivative of NCTC8325-4. Nevertheless, RN4220 was mutagenized via nitroguanoside to augment for mutants that would competently recognize plasmid DNA from E. coli [19, 29]. As a result, RN4220 carries 121 SNP's (eleven shared by NCTC8325-4), 4-large scale deletions, an insertion in agrA, and a nonsense mutation in hsdR (the major restriction endonuclease in S. aureus) (Nair et al., 2011) when compared to S. aureus NCTC8325. The agrA mutation present in RN4220 has been shown to lead to delayed activation of the agr quorum sensing system, which in turn, affects virulence factor production. Therefore, RN4220 is not suggested for the study of S. aureus virulence regulation or in vivo pathogenesis. As an alternative, RN4220 is used as a cloning intermediate when moving plasmid DNA from E. coli into S. aureus.
The eventual MSSA isolate broadly utilized in the laboratory is S. aureus Newman. Newman is not a derivative of NCTC8325, but was isolated as a substitute from a secondary osteomyelitis infection of a TB patient prior to 1952 (DUTHIE and LORENZ, 1952). Like NCTC8325, and all of its derivatives, Newman has distinctive features, including: the overproduction of toxins resulting from the constitutive expression of saeRS [1] as well as mutations in genes encoding for the virulence proteins FnbA and FnbB that limits their surface localization [18]. Furthermore, the S. aureus Newman genome contains four prophages (ΦNM1-4), three of which (ΦNM1, 2, and 4) are known to replicate during in vitro culture and animal infections [4]. In spite of these observations, Newman is generally utilized for virulence studies of S. aureus.
The most frequently studied MRSA isolates include: COL, Mu50, N315, JH1, JH9, UAMS-1, MRSA252, LAC, SF8300, FPR3757, TCH1516, and MW2 (Table 1). The entire MRSA isolates are commonly divided into hospital and community acquired strains (HA and CA-MRSA) based on the origin of the disease-causing isolate. Interestingly, this distinction has also come to reflect historical development of S. aureus pathogenesis and the emergence of a new MRSA-epidemic. HA-MRSA was first isolated in 1961 and quickly became the dominant hospital acquired infection following the widespread introduction of methicillin treatment in hospitals [16]. The most important HA-MRSA strains studied in the laboratory are COL [12, 44], Mu50 which is intermediate-vancomycin resistance strain [20, 30], N315 [30], MRSA252 [21], UAMS-1 [6], JH1, and JH9 [47]. These strains stand for the historical evolution of HA-MRSA as well as its geographic variety, and encompass two main pulse field gel electrophoresis (PFGE) subtypes, USA100 and USA200 [49]. HA-MRSA conventionally causes septicemia, osteomyelitis and surgical site infections (SSIs) in hospital patients.
CA-MRSA, on the other hand, was originally isolated in the late 1990's when healthy persons within the community, who had had no new contact with a hospital setting, contracted fatal S. aureus infections (CA-MRSA) [49]. It was later on determined that these isolates were different from their HA-MRSA foils in that they typically cause skin and soft tissue infections (SSTI's), sporadically cause necrotizing pneumonia, and can readily infect healthy persons [35]. The majority of frequently utilized CA-MRSA strains studied in the laboratory are SF8300 [7], LAC [28], FPR3757 [9], TCH1516 [17], and MW2 [3]. SF8300, LAC, FPR3757, and TCH1516 all fit in to the prevailing CA-MRSA PFGE subtype, USA300, whilst MW2 belongs to the second most common CA-MRSA PFGE subtype, USA400 [49]. Supplementary CA-MRSA lineages are newly recognized in the literature, but laboratory protocol characterizing these strains has merely recently been undertaken (e.g. ST239).
Although the inconsistency in pathogenesis between HA and CA-MRSA isolates implies detectable genetic distinctions between these two MRSA sub-groups, CA-MRSA isolates demonstrate a large amount of heterogeneity world-wide, making it hard to propose any single molecular test for identification of CA-MRSA isolates. Additionally, CA-MRSA strains have begun to swap HA-MRSA strains in many hospitals around the world, confusing the conventional classification of MRSA isolates into the HA and CA categories [23, 25, 36, 43]. Therefore, MRSA isolates have more recently been categorized into HA and CA sub-categories based on sequence typing (i.e. phylogeny) and pulse-field gel electrophoresis, of digested genomic DNA [14, 35]. Nevertheless, some uncomplicated molecular characteristics do separate CA-MRSA isolates from their HA-MRSA counterparts, notwithstanding these characteristics are not shared by every CA-MRSA strain (Table 2) [32, 49].
Growth of S. aureus in tuna was then monitored using Baird Parker agar; simultaneously, aerobic plate counts, enterotoxin production, and sensory profile (color and odor) were determined. The results showed that the time for a 3-log CFU/g increase was >20 h at 21°C and 8 to 12 h at 27°C for albacore, with toxin production observed at 14 to 16 h at 21°C and at 8 h at 27°C. A 3-log CFU/g increase for skipjack occurred at 22 to 24 h at 21°C and at 10 to 14 h at 27°C. The toxin production in skipjack started at 20 to 22 h at 21°C and at 8 to 10 h at 27°C. Toxin production was observed before a 3-log increase was achieved in albacore samples at 21°C. Under all conditions, toxins were detected when the cell density of S. aureus was 6 log CFU/g [53].
Reference materials are crucial to enforce legislation and to implement and safeguard reliable measurements. First, a feasibility study revealed a suitable processing procedure for cheese powders: the blank material was prepared by cutting, grinding, freeze-drying and milling. For the spiked material, cheese-water slurry was spiked with Staphylococcal enterotoxin solution, freeze-dried and diluted with blank material to the desired Staphylococcal enterotoxin concentration [55].
2.1. Fundamental Procedure 1: Growing S. aureus on Solid Media
S. aureus forms small, shiny, gold colonies on most solid media after 1 day of growth at 37°C. The size and color of the colonies is media and strain dependent (media containing low levels of glycolytic carbon sources will produce colonies that is less pigmented as are some strains harboring specific mutations that affect pigmentation). Numerous special media have been described for growing S. aureus including: Tryptic Soy Agar (TSA), Brain Heart Infusion (BHI) Agar, Mueller-Hinton Agar (MHA), Todd Hewitt Agar (THA), Luria-Bertani (LB) Agar, and Blood Agar. BHI, TSA, THA, MHA, LB Agar are all very rich media and will permit for vigorous growth of S. aureus. Blood agar is also a rich media but permits for the supplementary examination of hemolysis. The majority strains of S. aureus are β-hemolytic (i.e. produce complete lysis of red blood cells resulting in a zone of clearing) as a result of alpha toxin production. S. aureus colonies grown on blood agar are small, shiny, pigmented (to varying degrees depending on the glucose content of the blood agar used and the strain of S. aureus), and surrounded by a zone of clearing. Antibiotics can be added to the media as necessary according to the concentrations outlined in Table 2. It should be well-known that USA300 isolates enclose plasmids carrying an erythromycin and tetracycline resistance cassettes and therefore genetic management of USA300 involving antibiotic selection is limited.
2.2. Fundamental Procedure 2: Testing for Methicillin Resistance in S. aureus
As mentioned in tactical preparation, S. aureus resistance to penicillinase-stable penicillins (i.e. methicillin, oxacillin, nafcilin, etc.) is encoded by the mecA gene located on the SCCmec cassette [50]. The most accurate method for identifying methicillin-resistant Staphylococcus aureus (MRSA) is to test for the presence of the mecA gene by PCR [51]. Nevertheless, phenotypic confirmation of methicillin resistance is highly recommended regardless of the outcome of the PCR reaction as strains carrying the mecA gene can exhibit susceptibility to methicillin [2, 5]. The primary methods for demonstrating S. aureus methicillin resistance are the cefoxitin disk screen test, the latex agglutination test for PBP2a (the enzyme encoded by mecA), and the Oxacillin-salt agar screen [45]. While the latex agglutination test for PBP2a has been shown to be the most accurate method for determining S. aureus resistance to methicillin, the Oxacillin-salt agar screen is the simplest method and is still very accurate (99% sensitivity and 98.1% specificity) [45]. According to the Clinical and Laboratory Standards Institute (CLSI) S. aureus strains exhibiting growth on MHA supplemented with 2% NaCl and ≥ 4 μg/ml Oxacillin are considered methicillin-resistant while S. aureus strains only exhibiting growth on MHA supplemented with 2% NaCl when Oxacillin-levels are ≤ 2 μg/ml are considered methicillin-susceptible. Further information about the assay described below can be found in the Performance Standards for Antimicrobial Susceptibility Testing; Seventeenth Informational Supplement published by the CLSI (document M100-S17).
2.3. Fundamental Procedure 3: Growing of S. aureus in Liquid Media
S. aureus grows rapidly in broth culture at 37°C with aeration. As such, overnight cultures of S. aureus are typically started at the end of the day just prior to leaving the lab using either an isolated colony from an agar plate (Basic Protocol 1), a second broth culture, or a frozen stock (Basic Protocol 4). Quite a few different media have been described for growing S. aureus in broth culture, including: Brain Heart Infusion (BHI), Tryptic Soy Broth (TSB), Todd Hewitt Broth (THB), Luria-Bertani (LB) Broth, and Chemically Defined Media (CDM). There is no specific advantage or disadvantage to using BHI, TSB, THB, or LB; all are rich media and will allow for S. aureus to grow to a high concentration overnight. CDM, on the other hand, is commonly utilized for growth experiments relating the restriction or adding of specific nutrients. Not considering of the media, it should be noted that S. aureus is auxotrophic for Arginine [13] and Proline [31], that S. aureus cannot use inorganic sulfur sources (e.g. sulfate or sulfite) [48], and that S. aureus exhibits a preference for glycolytic carbon sources during aerobic growth [46]. Frequent passaging of S. aureus in liquid media can result in the accumulation of mutations and thus, should be avoided. Overnight cultures of S. aureus will exhibit gold pigmentation, although the extent of pigmentation is media and strain dependent.
2.4. Fundamental Procedure 4: Preparing Frozen Stocks of S. aureus
Durable storage of S. aureus should arise at −80°C to stop increase of mutations. To prepare a frozen stock of S. aureus add an aliquot of an overnight culture (Basic Protocol 3) to sterile DMSO as described below. The addition of DMSO thwart absolute freezing of the cells and hence limits cellular harm as a result of the transition to −80°C. Sterile glycerol (final concentration 25–50% once mixed with bacteria) is frequently used as an alternative to DMSO. There is no separate benefit to use one agent over the other except for that DMSO thaws more slowly than glycerol, and consequently preserves a low culture temperature for a longer period of time if freezer malfunction should occur.
3. How to Culture Staphylococcus aureus on Liquid and Solid Media
3.1. Brain Heart Infusion (BHI) Agar Plates
About 52g of BD BBL™ Brain Heart Infusion Agar should be added to distilled water in a 2L flask. Alternatively, 40g of BBL™ Tryptic Soy Agar or BBL™ Luria-Bertani Agar can be similarly utilized. Add a stir bar to the flask and then place the flask on a stir plate to mix. Bring volume up to 1L using distilled water. Loosely cover the top of the flask with aluminum foil and then autoclave the media on liquid cycle (use 30 minute cycle to prevent carmelization of glucose). Once sterilized, place the flask on a stir plate and stir the media at a low setting for ~45 minutes or until the media has cooled to 65°C. Add antibiotics as necessary and then pour the agar into sterile petri plates (~20–25ml/plate). Allow agar to solidify at room temperature for at least 1 day (minimizes condensation) then store up the plates, top down, in plastic bags at 4°C.
3.2. Brain Heart Infusion (BHI) Broth
About 37g of BD BBL™ Brain Heart Infusion should be added to distilled water in a 2L flask. On the other hand, 30g of BBL™ Tryptic Soy Agar, 25g of BBL™ Luria-Bertani Agar or 30 g of BBL™ Todd-Hewitt Broth can be equally utilized. Add a stir bar to the bottle and put the bottle on a stir plate to mix. Bring volume up to 1L using distilled water. Once the powder is totally dissolved, remove the stir bar, loosely cap the bottle, and autoclave the media on liquid cycle (do not tightly screw on the bottle cap or moreover the bottle will become highly pressurized in the autoclave; use 30 minute cycle to prevent glucose carmelization). Once sterilized, the broth may be stored at room temperature or 4°C.
3.3. Mueller-Hinton Agar Plates Supplemented with 2% NaCl and Oxacillin (MHA-Salt Plates)
About 38g of BD BBL™ Mueller-Hinton II Agar and 20g NaCl should be added to distilled water in a 1L bottle. Add a stir bar to the flask and then place the flask on a stir plate to mix. Bring volume up to 1L using distilled water. Loosely cover the top of the flask with aluminum foil and then autoclave the media on liquid cycle (use 30 minute cycle). Once sterilized, put the flask on a stir plate and stir the media at a low setting for ~45 minutes or until the media has cooled to 65°C. Add 2μg/ml and 4μg/ml Oxacillin as necessary and then pour the agar into sterile petri plates (~20–25ml/plate). Allow agar to solidify at room temperature for at least 1 day which minimizes condensation then store the plates, top down, in plastic bags at 4°C.
3.4. Chemically Distinct Culture Media
Make the solutions and sterilize (the amino acids and bases are made and stored separately to maximize the longevity of storage). Once cooled, all solutions should be stored at 4°C. Place a stir bar into a 250ml beaker and place the beaker onto a stir plate. Add 10mls of the salt solution, 1ml of each amino acid, 1ml of each base, 0.1ml of the vitamin solution, and 0.1ml of the trace elements solution to the beaker while slowly stirring the mixture. Add carbon sources as desired and then adjust the pH of the media to 7.4 using 10M NaOH. Bring the final volume of the media up to 100ml using distilled water and then filter sterilize the media.
3.5. Sterile DMSO
Sterilize 100% DMSO using a syringe filter (0.2μm Nylon syringe filter attached to a 10ml syringe). The DMSO should not be sterilized using a nitrocellulose filter for the reason that it will dissolve once it is exposed to the solvent.
4. Conclusion
Appropriate growth and maintenance of S. aureus in the laboratory is not difficult. Inocula need not be large in order to generate robust growth, and chances for contamination are rare given the short incubation times/fast growth rate of S. aureus cultures. Additionally, contaminants are simplyvisible from S. aureus by several easily testable phenotypes, all of which S. aureus is usually positive for, including: gold pigmentation, coagulase activity, β-hemolysis, catalase activity, and growth on Mannitol salt agar. Repeated route of S. aureus in the laboratory is not recommended as it may result in the amassing of unwanted mutations. For growth on BHI agar, S. aureus should appear as small, shiny, gold colonies after 12–24 hrs of growth at 37°C. Similarly, overnight cultures of S. aureus in BHI broth should show golden and highly turbid. S. aureus is a quick growing microorganism. Liquid cultures usually need just 12–24hrs to attain a high density while most strains of S. aureus will grow to be visible as colonies on a plate after 12–36hrs of growth at 37°C.
References
- Adhikari, R. P., Novick, R. P. (2008). Regulatory organization of the staphylococcal sae locus. Microbiology, 154: 949–959.
- Araj, G. F., Talhouk, R. S., Simaan, C. J., Maasad, M. J. (1999). Discrepancies between mec A PCR and conventional tests used for detection of methicillin resistant Staphylococcus aureus. International Journal of Antimicrobial Agents, 11: 47–52.
- Baba, T., Takeuchi, F., Kuroda, M., Yuzawa, H., Aoki, K. I., Oguchi, A., Nagai, Y., Iwama, N., Asano, K., Naimi, T., et al. (2002). Genome and virulence determinants of high virulence community-acquired MRSA. The Lancet, 359: 1819–1827.
- Bae, T., Baba, T., Hiramatsu, K., Schneewind, O. (2006). Prophages of Staphylococcus aureus Newman and their contribution to virulence. Molecular Microbiology, 62: 1035–1047.
- Bignardi, G. E., Woodford, N., Chapman, A., Johnson, A. P., Speller, D. (1996). Detection of the mec-A gene and phenotypic detection of resistance in Staphylococcus aureus isolates with borderline or low-level methicillin resistance. The Journal of antimicrobial chemotherapy, 37: 53–63.
- Cassat J (2006). Transcriptional profiling of a Staphylococcus aureus clinical isolate and its isogenic agr and sarA mutants reveals global differences in comparison to the laboratory strain RN6390. Microbiology, 152: 3075–3090.
- Charlebois, E. D., Bangsberg, D. R., Moss, N. J., Moore, M. R., Moss, A. R., Chambers, H. F., Perdreau-Remington, F. (2002). Population-based community prevalence of methicillin-resistant Staphylococcus aureus in the urban poor of San Francisco. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America. 34: 425–433.
- Climo, M. W. (2009). Decreasing MRSA infections: an end met by unclear means. JAMA: the journal of the American Medical Association,301: 772–773.
- Diep, B. A., Gill, S. R., Chang, R. F., Phan, T. H., Chen, J. H., Davidson, M. G., Lin, F., Lin, J., Carleton, H. A., Mongodin, E. F., et al(2006). Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. The Lancet, 367: 731–739.
- Dinges, M. M., Orwin, P. M., Schlievert, P. M. (2000). Exotoxins of Staphylococcus aureus. Clinical microbiology reviews. 13: 16–34.
- Duthie, E. S., Lorenz, L. L. (1952). Staphylococcal coagulase; mode of action and antigenicity. Journal of general microbiology. 6: 95–107.
- Dyke, K. (1969). Penicillinase production and intrinsic resistance to penicillins in methicillin-resistant cultures of Staphylococcus aureus. Journal of medical microbiology, 2: 261–278.
- Emmett, M., Kloos, W. E. (1979). The nature of arginine auxotrophy in cutaneous populations of staphylococci. Journal of general microbiology, 110: 305–314.
- Enright, M. C., Day, N. P., Davies, C. E., Peacock, S. J., Spratt, B. G. (2000). Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. Journal of clinical microbiology, 38: 1008–1015.
- Foster, T. J. (2005). Immune evasion by staphylococci. Nature Reviews Microbiology, 3: 948–958.
- Foster, T. J., Höök, M. (1998). Surface protein adhesins of Staphylococcus aureus. Trends in microbiology. 6: 484–488. Available at: http://ac.els-cdn.com/S0966842X98014000/1-s2.0-S0966842X98014000main.pdf?_tid=b5b27a903951069fdf4d8b989a649ca9&acdnat=1342717155_1dddd7a81cba19ba70448214d62dd903.
- Gonzalez, B. E. (2005). Severe Staphylococcal Sepsis in Adolescents in the Era of Community-Acquired Methicillin-Resistant Staphylococcus aureus. Pediatrics.115: 642–648.
- Grundmeier M, Hussain M, Becker P, Heilmann C, Peters G, Sinha B (2004). Truncation of Fibronectin-Binding Proteins in Staphylococcus aureus Strain Newman Leads to Deficient Adherence and Host Cell Invasion Due to Loss of the Cell Wall Anchor Function. Infection and immunity, 72: 7155–7163.
- Herbert, S., Ziebandt, A. K., Ohlsen, K., Schäfer, T., Hecker, M., Albrecht, D., Novick, R., Gotz, F. (2010). Repair of Global Regulators in Staphylococcus aureus 8325 and Comparative Analysis with Other Clinical Isolates. Infection and immunity, 78: 2877–2889.
- Hiramatsu, K., Hanaki, H., Ino, T., Yabuta, K., Oguri, T., Tenover, F. C. (1997). Methicillin-resistant Staphylococcus aureus clinical strain with reduced vancomycin susceptibility. The Journal of antimicrobial chemotherapy, 40: 135–136.
- Holden, M. T. G., Feil, E. J., Lindsay, J. A., Peacock, S. J., Day, N. P. J.., Enright, M. C., Foster, T. J., Moore, C. E., Hurst, L., Atkin, R. et al. (2004). Complete genomes of two clinical Staphylococcus aureus strains: evidence for the rapid evolution of virulence and drug resistance. Proceedings of the National Academy of Sciences of the United States of America, 101: 9786–9791.
- Horsburgh, M. J., Aish, J. L., White, I. J., Shaw, L., Lithgow, J. K., Foster, S. J. B.(2002). Modulates Virulence Determinant Expression and Stress Resistance: Characterization of a Functional rsbU Strain Derived from Staphylococcus aureus 8325-4. Journal of Bacteriology, 184: 5457–5467.
- Hultén, K. G., Kaplan, S. L., Lamberth, L. B., Slimp, K., Hammerman, W. A., Carrillo-Marquez, M., Starke, J. R., Versalovic, J., Mason, E. O. (2010). Hospital-Acquired Staphylococcus aureusInfections at Texas Children's Hospital, 2001–2007. Infection control and hospital epidemiology:the official journal of the Society of Hospital Epidemiologists of America, 31: 183–190.
- International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC, 2009) Classification of Staphylococcal Cassette Chromosome mec (SCCmec): Guidelines for Reporting Novel SCCmec Elements. Antimicrobial Agents and Chemotherapy, 53: 4961–4967.
- Jenkins, T. C., McCollister, B. D., Sharma, R., McFann, K. K., Madinger, N. E., Barron, M., Bessesen, M., Price, C. S., Burman, W. J. (2009). Epidemiology of Healthcare-Associated Bloodstream Infection Caused by USA300 Strains of Methicillin-Resistant Staphylococcus aureusin 3 Affiliated Hospitals. Infection control and hospital epidemiology:the official journal of the Society of Hospital Epidemiologists of America, 30: 233–241.
- Jevons, M. P. (1961). "Celbenin" - Resistant Staphylococci. British Medical Journal, 124–125.
- Katzif, S., Lee, E. H., Law, A. B., Tzeng, Y. L., Shafer, W. M. (2005). CspA regulates pigment production in Staphylococcus aureus through a SigB-dependent mechanism. Journal of Bacteriology, 187: 8181–8184.
- Kennedy, A. D., Otto, M., Braughton, K. R., Whitney, A. R., Chen, L., Mathema, B., Mediavilla, J. R., Byrne, K. A., Parkins, L. D., Tenover, F. C., et al.(2008). Epidemic community-associated methicillin-resistant Staphylococcus aureus: recent clonal expansion and diversification. Proceedings of the National Academy of Sciences, 105: 1327–1332.
- Kreiswirth, B. N., Löfdahl, S., Betley, M. J., O'Reilly, M., Schlievert, P. M., Bergdoll, M. S., Novick, R. P. (1983). The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature, 305: 709–712.
- Kuroda, M, Ohta T, Uchiyama I, Baba T, Yuzawa, H., Kobayashi, I., Cui, L., Oguchi, A., Aoki, K. I., Nagai, Y., et al.(2001). Whole genome sequencing of meticillin-resistant Staphylococcus aureus. The Lancet, 357: 1225–1240.
- Li, C., Sun F, Cho H, Yelavarthi V, Sohn C, He C, Schneewind O, Bae T (2010). CcpA Mediates ProlineAuxotrophy and Is Required for Staphylococcus aureus Pathogenesis. Journal of Bacteriology, 192: 3883–3892.
- Li, M., Du, X., Villaruz, A. E., Diep, B. A., Wang, D., Song, Y., Tian, Y., Hu, J, Yu, F., Lu, Y., et al(2012). MRSA epidemic linked to a quickly spreading colonization and virulence determinant. Nature Medicine, 18: 816–819.
- Liu, G. Y. (2005). Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. Journal of Experimental Medicine, 202: 209–215.
- McDougal, L. K., Steward, C. D, Killgore, G. E., Chaitram, J. M., McAllister, S. K., Tenover, F. C. (2003). Pulsed-Field Gel Electrophoresis Typing of Oxacillin-Resistant Staphylococcus aureus Isolates from the United States: Establishing a National Database. Journal of clinical microbiology, 41: 5113–5120.
- Millar, B. C., Loughrey, A., Elborn, J. S., Moore, J. E. (2007). Proposed definitions of community-associated meticillin-resistant Staphylococcus aureus (CA-MRSA) Journal of Hospital Infection, 67: 109–113.
- Moore, C. L., Hingwe, A., Donabedian, S. M., Perri, M. B., Davis, S. L., Haque, N. Z., Reyes, K., Vager, D., Zervos, M. J. (2009). Comparative evaluation of epidemiology and outcomes of methicillin-resistant Staphylococcus aureus (MRSA) USA300 infections causing community- and healthcare-associated infections. International Journal of Antimicrobial Agents, 34: 148–155.
- Nair, D., Memmi, G., Hernandez, D., Bard, J., Beaume, M., Gill, S., François, P., Cheung, A. L. (2011). Whole-Genome Sequencing of Staphylococcus aureus Strain RN4220, a Key Laboratory Strain Used in Virulence Research, Identifies Mutations That Affect Not Only Virulence Factors but Also the Fitness of the Strain. Journal of Bacteriology, 193: 2332–2335.
- Novick, R. P. (1967). Properties of a cryptic high-frequency transducing phage in Staphylococcus aureus. Virology, 33: 155–166.
- Novick, R. P., Richmond, M. H. (1965). Nature And Interactions of The Genetic Elements Governing Penicillinase Synthesis In Staphylococcus aureus.Journal of Bacteriology,90: 467–480.
- Ogston, A. (1881). Report upon Micro-Organisms in Surgical Diseases. British Medical Journal, 1: 369–375.
- Pelz, A., Wieland, K. P.., Putzbach, K., Hentschel, P., Albert, K., Götz, F. (2005). Structure and biosynthesis of staphyloxanthin from Staphylococcus aureus. The Journal of biological chemistry, 280: 32493–32498.
- Peng, H. L., Novick, R. P., Kreiswirth, B., Kornblum, J., Schlievert, P. (1988). Cloning, characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus aureus. Journal of Bacteriology, 170: 4365–4372.
- Popovich, K. J., Weinstein, R. A., Hota, B. (2008). Are Community-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) Strains Replacing Traditional Nosocomial MRSA Strains? Clinical infectious diseases: an official publication of the Infectious Diseases Society of America. 46: 787–794.
- Sabath, L. D., Wallace, S. J., Gerstein, D. A. (1972). Suppression of Intrinsic Resistance to Methicillin and Other Penicillins in Staphylococcus aureus. Antimicrobial Agents and Chemotherapy, 2: 350–355.
- Sakoulas, G., Gold, H. S., Venkataraman, L., DeGirolami, P. C., Eliopoulos, G. M., Qian, Q. (2001). Methicillin-Resistant Staphylococcus aureus: Comparison of Susceptibility Testing Methods and Analysis of mecA-Positive Susceptible Strains. Journal of clinical microbiology, 39: 3946–3951.
- Seidl, K., Müller, S., Francois, P., Kriebitzsch, C., Schrenzel, J., Engelmann, S., Bischoff, M., Berger-Bächi, B. (2009). Effect of a glucose impulse on the CcpAregulon in Staphylococcus aureus. BMC Microbiology, 9: 95.
- Sieradzki, K., Leski, T., Dick, J., Borio, L., Tomasz, A. (2003). Evolution of a vancomycin-intermediate Staphylococcus aureus strain in vivo: multiple changes in the antibiotic resistance phenotypes of a single lineage of methicillin-resistant S. aureus under the impact of antibiotics administered for chemotherapy. Journal of clinical microbiology, 41: 1687–1693.
- Soutourina, O., Poupel, O., Coppée, J. Y., Danchin, A., Msadek, T., Martin-Verstraete, I. (2009). CymR, the master regulator of cysteine metabolism in Staphylococcus aureus, controls host sulphur source utilization and plays a role in biofilm formation. Molecular Microbiology, 73: 194–211.
- Thurlow, L. R., Joshi, G. S., Richardson, A. R. (2012). Virulence strategies of the dominant USA300 lineage of community-associated methicillin-resistant Staphylococcus aureus (CAMRSA) FEMS Immunology & Medical Microbiology, 65: 5–22.
- Ubukata, K., Nonoguchi, R., Matsuhashi, M., Konno, M. (1989). Expression and inducibility in Staphylococcus aureus of the mecA gene, which encodes a methicillin-resistant S. aureus-specific penicillin-binding protein. Journal of Bacteriology, 171: 2882–2885.
- Unal, S., Hoskins, J., Flokowitsch, J. E., Wu, C. Y., Preston, D. A., Skatrud, P. L. (1992). Detection of methicillin-resistant staphylococci by using the polymerase chain reaction. Journal of clinical microbiology, 30: 1685–1691.
- Weber, J. T. (2005). Community-associated methicillin-resistant Staphylococcus aureus. Clinical Infectious Diseases, 41 (Suppl 4): S269–72.
- Kataoka, A. I., Enache, E., Napier, C., Hayman, M., Weddig, L. (2016). Effect of Storage Temperature on the Outgrowth and Toxin Production of Staphylococcus aureus in Freeze-Thawed Precooked Tuna Mea. Journal of Food Protection. 4 (1): 524-686.
- Zeleny, R., Emteborg, H., Charoud-Got, J., Schimmel, H., Nia, Y., Mutel, I., Ostyn, A., Herbin, S., Hennekinne, J. A. (2015 Development of a reference material for Staphylococcus aureus enterotoxin A in cheese: Feasibility study, processing, homogeneity and stability assessment. Food Chemistry. 168 (1): 241–246.



