
American Journal of Microbiology and Immunology, Vol. 1, No. 2, September 2016 Publish Date: Sep. 23, 2016 Pages: 26-31
Evaluation of the Larvicidal Activity of Bacillus Thuringiensis Isolated from Soil Samples Against Aedes aegypti
Bello A.*, Machido D. A., Inabo H. I.
Department of Microbiology, Faculty of Science, Ahmadu Bello University, Zaria, Nigeria
Abstract
Application of chemical insecticides poses a wide range of problems in the environment. In addition to this, these chemicals are non specific and are implicated as carcinogens. All these necessitated the search for an eco-friendly biological alternative for the control of insect pests and vectors in environment such as the use of Bacillus thuringiensis known to habour insecticidal crystals. Bioassay to assess the insecticidal activity of the isolates was carried out on Aedes aegypti larvae using 3 different concentrations of spore crystal mixture (100, 75 and 50 ppm) alongside one control. In each case, 10 larvae were exposed in triplicates for Aedes aegypti. The mortality range of Aedes aegyti was between 40.00% and 100.00%. The concentrations of the spore crystal mixture which kill 50% of the exposed populations in standard bioassays (LC50) were determined by probit analyses. The LC50 using was between 118.03ppm and 55.79ppm. The results of this research shows that the isolates from the soils in Zaria can serve as biocontrol agent for the control of mosquito by targeting their larvae stage hence controlling the diseases they spread.
Keywords
Larvicidal, Bacillus Thuringiensis, Aedes Aegyti, Biocontrol, LC50
Received: August 9, 2016
Accepted: September 7, 2016
Published online: September 23, 2016
@ 2016 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY license. http://creativecommons.org/licenses/by/4.0/
Contents
1. Introduction 2. Materials and Methods 2.1. Collection of Samples 2.2. Isolation Procedure 2.3. Characterization of Bacillus thuringiensis 2.4. Preparation of Spore Crystal Mixture 2.5. Collection and Counting of Mosquito Larva 2.6. Determination of the Larvicidal Activity of Bacillus thuringiensis Against Aedes aegypti Larvae 2.7. Data Analysis 3. Results and Discussion 4. Conclusion
1. Introduction
Bacillus thuringiensis belongs to the family of the Bacillaceae; which Taxonomists consider as sub-species of Bacillus cereus because they are closely related [9, 14, 16]. According to [18], the genotypic and phenotypic characteristics of Bacillus thuringiensis are very similar to Bacillus cereus; the only difference between these two species being the formation of large proteinaceous parasporal inclusions observed in Bacillus thuringiensis.
Around the world, the medical and economic burden caused by vector-borne diseases continues to grow as current control measures fail to cope. There is an urgent need to identify new control strategies that will remain effective, even in the face of growing insecticide and drug resistance [1].
Repetitive use of man-made insecticides for mosquito control disrupts natural biological control systems and lead to reappearance of mosquito populations. It also resulted in the development of resistance, detrimental effects on non-target organisms and human health problems and subsequently this initiated a search for alternative control measures [11].
Mosquitoes, which are vectors for many diseases belong to the family Culicidae in the insect order of true-flies or two-winged flies called Diptera. During these blood meals, a female mosquito transmits agents of diseases to man.
Aedes aegypti (Linnaeus) belongs to the family Culicidae and the order Diptera. It is the vector of diseases such as dengue fever, chikungunya and yellow fever dengue hemorrhagic fever, other viral diseases, lymphatic filariasis. It is also been implicated in the transmission of Zika virus which is suspected of causing a neurological disorder called the Guillain Barre syndrome. The female A. aegypti preferably lay eggs in artificial collections of water. The hatched larvae undergo growth and metamorphosis.
Insect pests of crops and forest plants and vectors of disease of human beings and other animals are serious threat for agriculture and public health. Worldwide, about US $8000 billion is spent for insecticides and estimates reveal that US $2700 can be substituted by the biopesticide B. thuringiensis. Besides exorbitant cost, and resistance and resurgence of the different pests, the chemical pesticides are the single main cause of health and environmental hazard. The situation demands the safer pesticides and biopesticides are the most desired alternatives. Bacteria, especially B. thuringiensis and B. sphaericus are the most potent and successful group of organisms for effective control of insect pests and vectors of diseases [8].
Owing to the non-specificity of chemical insecticides, both target and non target organisms are often impacted upon with undesirable consequences. As a result, organisms that play an important role in the environment could be detrimentally affected. Key insect pollinators and other resourceful life forms could be wiped out [5]. Both surface and underground water bodies could become heavily contaminated with insecticide residues. This could pose a serious threat to aquatic life forms. [10]. These problems are further compounded by exorbitant cost of procurement and almost total lack of proper knowledge of their application among the general populace [5].
B. thuringiensis has certain advantages for exploitation as biopesticide viz. B. thuringiensis can be used directly and as transgenic microbes and plants, being a prokaryote there is no dominant or recessive allele, highly vulnerable to genetic manipulation and the toxin gene is coded by single gene (monocistronic), B. thuringiensis is fermentation friendly and therefore commercially exploitable and it is host specific or has narrow host range [8].
The predominant type of δ-endotoxins found in B. thuringiensis is the Cry proteins, a crystal protein encoded by Cry genes. The accumulation of Cry protein in a mother cell can make up 20-30% of the dry weight of the sporulated cells [3, 4].
Each Crystal protein has its own insecticidal spectrum. The crystal protein of B. thuringiensis acts as a protoxins, for this protoxins to become active; a suspectible insect must ingest them. After being ingested, the crystals are solubilized in the alkaline environment in the insect midgut (pH>10). After solubilization, enzymes in midgut (proteases) convert the protoxins into active toxins. This active toxin then binds to specific receptors on the membranes of epithelial midgut cells; this interaction provides the insertion of the toxin into the lipid bilayer and formation of pores (0.5 to 1 nm). This pore formation leads to gut paralysis. Finally, insect larvae stop feeding and die from lethal septicemia [2, 12].
Present study was envisaged to isolate and identify the B. thuringiensis of indigenous soils of Zaria, Kaduna state, Nigeria and characterized the polymorphic crystal producing strains, which was exploited for biological control of mosquito insect-pests which are disease vectors.
2. Materials and Methods
2.1. Collection of Samples
Samples of organic matter-rich soil were collected from various sites in Zaria. From each location, samples of about l0g were collected from five spots. Samples were taken from one inch below the surface by scrapping off surface material with a spatula. The samples were brought to the laboratory and stored under appropriate storage conditions.
2.2. Isolation Procedure
Bacillus thurigiensis were isolated according to the method described by Travers et al. (1987). For each sample, 0.5 g soil was added in 10 ml of LB medium (Tryptone 10g /L, yeast extract 5g/L, NaCl 5g/L) to which 0.25M sodium acetate was added and incubated in shaking incubator at 30°C and 250 rpm for 4 hours. From each sample 2ml was taken and heat shocked in a water bath at 80°C for 20 minutes. Serial dilutions of treated samples were prepared and spread on T3 agar ((Tryptone 3g/L, Yeast extract 1.5g/L, Tryptose 2g/L, MnCl2 0.005g/L, Sodium phosphate 0.05M and Agar15g/L) and incubated for 2days at 26°C. Colonies with B. thuringiensis like morphology (entire margin, off white color, dry and rich growth of colony) were picked up at random and purified by streaking them on LB agar plates (Tryptone 3g/L, Yeast extract 1.5g/L, Tryptose 2g/L, MnCl2 0.005g/L, NaH2 PO4 6.9g/L, Na2HPO4 8.9g/L and Agar15g/L).
2.3. Characterization of Bacillus thuringiensis
The suspected isolates of Bacillus thuringiensis were identified by following the diagnostic plan specified in Bergey's Manual of Determinative Bacteriology [15]. The isolate were characterized using Gram reaction and endospore staining, motility, growth above 45 °C, catalase activity, Voges Proskauer test and confirmed using MicrogenTM Bacillus-ID.
2.4. Preparation of Spore Crystal Mixture
To obtain the spore-crystal mixtures, single colonies from overnight LB plates were inoculated into 10ml T3 sporulation medium (per litre 3g of tryptone, 2g of tryptose, 1.5g of yeast extract, 0.05m sodium phosphate (pH 6.8) and 0.005g MnCl2) and cultured for 60 hours in a shaker incubator at 30°C. Spores and crystals were harvested by centrifugation at 7000g for 10 minutes and washed twice with distilled water, the spore crystal mixture were stored at low temperature until required [6, 13].
2.5. Collection and Counting of Mosquito Larva
The third instar larvae (L3) of Aedes aegypti were collected from discarded containers and stagnant water around the university. Early third instar (L3) Larvae of Aedes aegypti used in the bioassay were selected using a modified pasteur pipette and placing 10 early L3 larvae into each test tube and the volume was made up to 10 ml using distilled water.
2.6. Determination of the Larvicidal Activity of Bacillus thuringiensis Against Aedes aegypti Larvae
The mosquitocidal assay was performed using three concentrations of spores and crystal mixture (100ppm, 75ppm and 50ppm). Each concentration was replicated three times and one test tube with 10 (L3) larvae was used as the control. Mortality of the treated larvae were recorded after 24 hours [17].
2.7. Data Analysis
Results were presented in tables, graphs and charts where applicable. ANOVA was used to compare the mean mortality of the isolates at different concentration of spore crystal mixture of each of the isolates. Using probit analysis, the mean lethal concentration, LC50 was determined, for each of the isolates tested. Discussion and conclusions were made afterwards.
3. Results and Discussion
Chemical insecticides provide many benefits to food production and human health and have proven very effective at increasing agriculture and forestry productivities. However, they also pose some hazards such as contamination of water and food sources, poisoning of non-target fauna and flora, concentration in the food chain and selection of insect pest populations resistant to the chemical insecticides [20].
A total of 30 soil samples were collected. From these 30 soil samples, 12 Bacillus thuringiensis were isolated and characterized biochemically.
The bioassay for the larvicidal activity of the isolates on Aedes aegyti larvae using different concentrations of the spore crystal mixture shows that the higher the concentration of spore crystal mixture, the more the larvicidal activity.
Isolate L3 had the highest activity (100% mortality) at 100ppm concentration of spore crystal mixture, followed by L1 (93.33% mortality) at the same concentration. Isolate D2 shows the lowest activity (40.00%) at 100ppm concentration (Figure 1). The differences observed between the mean mortality of the isolates at different concentration of the spore crystal mixture were statistically significant (Table 1).
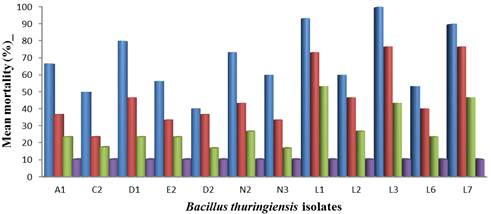
Figure 1. Mean mortality of Aedes aegypti to different concentrations of Bacillus thuringiensis spore crystal mixture after 24 hours of exposure.
Table 1. Mortality of Aedes aegypti larvae on exposure to various concentrations of twelve isolates of Bacillus thuringiensis.
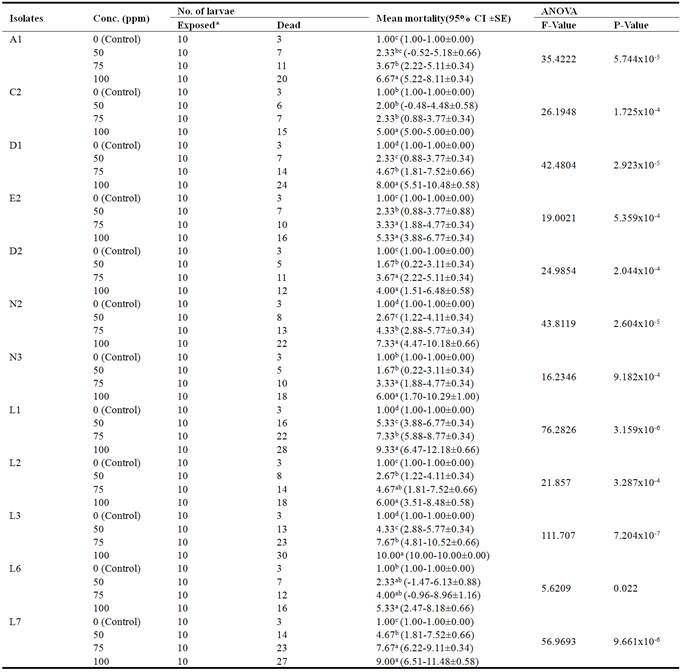
Means followed by the same superscript within the same isolate are not significantly different (P˃0.05).
* 10 larvae were exposed in triplicate giving a total of 30.
The probit analysis was used to assess the potency of the toxin on living organisms (as such it was used to assess the potency of the spore crystal mixture of the isolates on mosquito larvae) the probit table was used to get the Empirical Probit of kill for each of the concentrations using their corresponding mortality. The empirical probit of kill was plotted against the log of concentration to derive the regression equation. The coefficient of regression (R2) was also generated from the graph. The regression equation was used to calculate the median lethal concentration (LC50) which is the concentration that will kill 50% of the exposed population. The table below shows the median lethal concentration (LC50) of twelve isolates of Bacillus thuringiensis against Aedes aegypti larvae obtained by probit analysis.
The concentrations of the spore crystal mixture which kill 50% of the exposed populations in standard bioassays (LC50) were determined by probit analyses with triplicate bioassay. These concentrations range between 52.17ppm and 125.89ppm (Table 2).
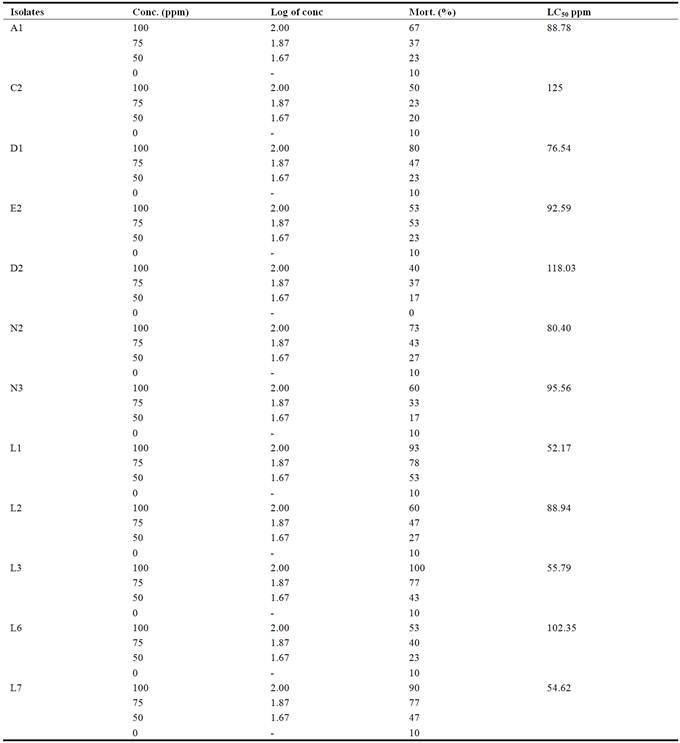
Conc. = concentration, Log of conc = logarithm of concentration, Mort. = mortality, LC50 % = Median Lethal concentration.
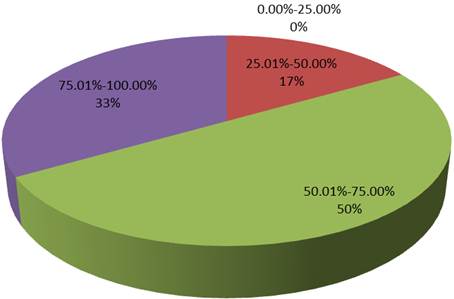
Based on the distribution of the insecticidal activity of the isolates Aedes aegypti; 0% of the isolates had activity between 0.00-25.00%, 17% had activity between 25.01%-50.00%, activity between 75.00%-100.0% was seen in 33% of the isolates, while activity between 50.01%-75.00% was seen in 50% (Figure 2).
4. Conclusion
The search for alternative biocontrol agents such as Bacillus thuringiensis-based biopesticides is increasingly attracting interest. This bacterium produces parasporal crystalline inclusions (Cry proteins) which is toxic to many important agricultural pests. The Cry proteins are encoded by cry genes and so far, many cry genes have been identified in different B. thuringiensis strain collections. Cry2 and cry4 are toxic to dipteran insects [7, 21].
Bioassay to screen for the larvicidal activity of the isolates against aegypti larvae showed that the isolates are promising biocontrol agents for the control of these mosquitoes. The bioassay also showed that most of the Bacillus thuringiensis isolates from the soil harbor dipteral-active cry hence they are good biocontrol agents for mosquitoes.
References
- Achs, J and Malaney, P. (2002). The economic and social burden of malaria. Nature, 15: 680-685.
- Adang, M. J. (1991). Bacillus thuringiensis insecticidal crystal proteins: gene structure, action and utilization. in Biotechnology for Biological Control of Pest and Vectors, edited by K. Maramorosh, (CRC Press, Boca Raton Fla, 1991), 3-24.
- Agaisse, H. and Lereclus, D. (1995). How does Bacillus thuringiensis produce so much insecticidal crystal protein?, Journal of Bacteriology, 177: 6027-6032.
- Baum, J. A, and Malvar, T. (1995). Regulation of insecticidal crystal protein production in Bacillus thuringiensis., Molecular Microbiology, 18: 1-12.
- Biswas, M. R. (1994). Agriculture and Environment: A Review. Ambio, 23 (3): 192-197.
- Carozzi, N. B., Kramer, V. C., Warren, G. W., Evola, S. and Koziel, M. G., (1991). Prediction of insecticidal activity of Bacillus thuringiensis strains by polymerase chain reaction product profiles. Applied environmental Microbiology, 57: 3057-3061.
- Çetinkaya, F. T. (2002). Isolation of Bacillus thuringiensis and Investigation of Its Crystal Protein Genes. An unpublished Thesis submitted to the Graduate School of Biotechnology and Bioengineering İzmir Institute of Technology, İzmir, Turkey.
- Chatterjee, S. N., Bhattacharya, T. Dangar, T. K. and Chandra, G. (2007). Ecology and diversity of Bacillus thuringiensis in soil environment. African Journal of Biotechnology, 6 (13): 1587-1591.
- Chen, M. L. and Tsen, H. Y. (2002). "Discrimination of Bacillus cereus and Bacillus thuringiensis with 16S rRNA and gyrB Gene Based PCR Primers and Sequencing of Their Annealing Sites,"Journal of Applied Microbiology, 92: 912-919.
- Cremlyn, R. J. (1991). Agrochemicals: Preparation and Mode of Action. John Wiley and Sons. London, New York. Pp. 157-216.
- Das, N. G., Goswami, D. and Rabhar, B. (2007). Preliminary evaluation of mosquito larvicidal efficacy of plant extracts. Journal of Vector Borne Diseases, 44: 145–148.
- Gill, S. S., Cowles, E. A. and Pietranto, P. V. (1992). The mode of action of Bacillus thuringiensis endotoxins., Annual Review of Entomology, 37: 615-636.
- Gorashi, N. E., Elshafie, H. A. F., Hamid, HA and Dirar, D. H. (2012). Characterization of sudan strain of Bacillus thuringiensis pathogenic to larvae of the house mosquito Culex quinquafasciatus. Agricultural Biological journal of North America, 3: 271.
- Helgason, E., Okstad, O. A., Caugant, D. A., Johansen, H. A., Fouet, A. and Mock, M. (2000). "Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis-one species on the Basis of Genetic Evidence," Applied and Environmental Microbiology, 66: 2627-2630.
- Holt, J. G., Kreig, N. R., Sneath, P. H. A., Staley, J. T., Willliams, S. T. (1994), Bergey’s Manual of Determinative Bacteriology, 9th Edition., Lippincott Williams and Wilkins, Baltimore.
- Leonard, C, Yahua, C. and Mahilion, J. (1997). "Diversity and Distribution of IS231, IS232, and IS240 among Bacillus cereus, Bacillus thuringiensis and Bacillus mycoides," Microbiology, 143: 2537-2547.
- Naiema, E. G., Hamadttu, A. F. E., Hamid, A. H. and Dirar, H. D. (2012). Characterization of Sudan strains of Bacillus thuringiensis pathogenic to the larvae of the house mosquito Culex quinquefasciatus. Agriculture and Biology Journal of North America, 3 (7): 271-279.
- Priest, F. G. (2000). "Biodiversity of the Entomopathogenic, Endospore-forming Bacteria'", in Entomopathogenic Bacteria: From laboratory to field application, edited by J. F. Charles, A. Delecluse and C. Nielsen-LeRoux (Kluwer Academic, Dordrecht), 1-22.
- Travers, R. S., Martin, P. A. W. and Reichelderfer, C. F. (1987). Selective process for efficient isolation of Bacillus spp., Applied Environmental Microbiology, 53: 1263-1266.
- Wojciech, J. J. and Korsten, L. (2002). Biological Control of Postharvest Diseases of Fruits. Annual Review of Phytopathology, 40: 411-441.
- Zeigler, D. R. (1999), Bacillus Genetic Stock Center Catalog of Strains, Seventh Edition, Part 2: Bacillus thuringiensis and Bacillus cereus, 37.
- Zhang, W. J, Jiang, F. B, and Ou, J. F. (2011). Global pesticide consumption and pollution: with China as a focus. Proceedings of the International Academy of Ecology and Environmental Sciences, 1 (2): 125-144.



