
American Journal of Microbiology and Immunology, Vol. 1, No. 2, September 2016 Publish Date: Aug. 25, 2016 Pages: 10-15
Effect of Temperature, pH and NaCl on the Isolates of Actinomycetes from Straw and Compost Samples from Savar, Dhaka, Bangladesh
Muhammad Ali Akond1, *, Mst Nusrat Jahan1, Nigar Sultana2,
Farhana Rahman3
1Department of Botany, Jahangirnagar University, Savar, Dhaka
2Legend College, Sobhanbag, Jaleshor, Savar, Dhaka
3Appartment No. 1405, Building No. 20, Japan Garden City, Mohammadpur, Dhaka
Abstract
The effect of physiological parameters viz., temperature, pH and NaCl on actinomycetes strains belonging to the genus Streptomyces (JUBM-35-NS-1) and Nocardia (JUBM-35-NS-2), isolated earlier from straw and compost materials used in commercial mushroom cultivation, was studied. Growth maxima were observed at 30°C and 35°C respectively in Streptomyces sp. and Nocardia sp. Arrest in growth for both isolates was found to be at 4°C and 50°C temperatures. The optimum growth was recorded at pH range of 6.0-6.5 for the isolate JUBM-35-NS-1 while it was at 6.5-8.0 for isolate JUBM-35-NS-2. Maximum growth was shown at NaCl concentration of 0% and 0.75% respectively by the isolate JUBM-35-NS-1 and JUBM-35-NS-2. The NaCl tolerance for growth of isolate JUBM-35-NS-1 was up to 8% and that for the isolate JUBM-35-NS-2 was up to 2.0%. A lower concentration of NaCl was found stimulatory with decline over 1.5% and complete cessation at 2.5% for the isolate JUBM-35-NS-2 (Nocardia sp.). The isolate JUBM-35-NS-1 (Streptomyces sp.) was found to be salt tolerant showing a stable growth from 0.5% to 8.0% with complete arrest of growth at 8.5% of NaCl concentration in culture medium.
Keywords
Actinomycetes, Growth, Temperature, pH, Salinity
Received:July 20, 2016
Accepted: August 9, 2016
Published online: August 25, 2016
@ 2016 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY license. http://creativecommons.org/licenses/by/4.0/
Contents
1. Introduction
Actinomycetes are Gram-positive bacteria frequently filamentous and sporulating, exhibiting a wide range of morphological forms from coccoid through fragmenting hypha to permanent and highly differentiated branched mycelium. Actinomycetes are present in various ecological habitats such as soil, fresh water, black water, lake, compost, sewage and marine environment [10]. Actinomycetes are the most fruitful source for production of bioactive secondary metabolites [5]. This is the largest group of microbes producing antifungal, antibacterial, and antiparasitic drugs. They also play very important role in producing wide range of bioactive compounds. Other biotechnological applications include the exploitation of their capacity of producing various enzymes which degrade complex organics such as cellulose, lignin, chitin, and proteins.
They play an important part in the cycle of life in nature by bringing about the decomposition of complex plant and animal residues and the liberation of a continuous stream of available elements, notably carbon and nitrogen, essential for fresh plant growth. Some of the biochemical activities of the actinomycetes are now being utilized for the large-scale production of chemical substances essential for public health [38]. The increased waste production is of great concern globally at different levels of population. Various alternatives are in exercise to address this crisis by elimination, purification and/or recycling. The modern concept of environmental management is based on the recycling of wastes. In this context, composting appears to be a safe form of treatment of some wastes and the reclamation of the nutrients [1]. Degradation of straw, the most abundant agricultural by-products with high amounts of cellulose and lignin, by a number of microbes would result in production of good quality compost containing higher amounts of total nitrogen, total potassium and total phosphorus content [36].
Microorganisms are specific to temperature, pH, and salinity for their growth, development and for the production of specific metabolites. For the production of bioactive secondary metabolites it is essential to maintain the optimum temperature, pH and salinity otherwise production could be failed or no growth also be observed. Isolation and identification of strains of Streptomyces sp. and Nocardia sp. were reported earlier from straw and compost samples [16] which were found to degrade rice straw. We report hereby the effect of temperature, pH and salinity on Streptomyces sp. (JUBM-35-NS-1) and Nocardia sp. (JUBM-35-NS-2) recovered previously.
2. Materials and Methods
2.1. Actinomycetes Isolates
The isolate JUBM-35-NS-1 previously identified as Streptomyces sp. was recovered from the straw sample [16]. During isolation chloramphenicol and nystatin was added to the medium and initially isolated at 45°C temperature in actinomycetes selective medium. The isolate JUBM-35-NS-2 identified as Nocardia sp. was recovered from compost sample at 45°C temperature in actinomycetes selective medium [16]. For the isolation from compost sample no antibiotic was used. In case of isolation of Streptomyces strain JUBM-35-NS-1 a small piece of straw was placed on selective agar plates; and for Nocardia strain JUBM-35-NS-2 a small amount of compost used in mushroom cultivation was directly inoculated by spreading over the solid surface of the selective medium. Subcultures were repeated until the colonies were found pure by visual and microscopic observation. Identification was made according to Bergey’s Manual of Determinative Bacteriology [13] based on morphological, physiological and biochemical characterization.
2.2. Effect of Temperature
0.1 ml of freshly developed seed cultures of the actinomycetes isolates in nutrient broth was inoculated into test tubes containing nutrient broth medium with the help of a micro pipette. Optical density (OD) was measured by a colorimeter at 600 nm light spectrum which was treated as initial density or density of control. Incubation was made at temperatures of 4°C, 25°C, 30°C, 37°C, 40°C, 45°C and 50°C for 48 hours in a shaker bath. After 48 hours, the optical density of each treatment was measured at 600 nm. Each treatment was replicated thrice. Initial OD was subtracted from that after 48 hours to calculate the growth obtained by each treatment.
2.3. Study on pH Effect
Nutrient broth was used to determine the effect of pH on the growth of actinomycetes isolates. Various pH values used were 4.0, 5.0, 5.5, 6.5, 7.0, 7.5, 8.0, 9.0, 10, and 11 adjusted with 1N HCl and 1N KOH solutions as per requirement. 0.1 ml seed cultures were inoculated in each tube with a micro pipette and the initial optical density was recorded. Incubation was made at 35°C temperature for 48 hours and the growth was measured by using optical density. Three tubes were incubated for each pH treatment.
2.4. Effect of NaCl
Nutrient broth in test tubes with NaCl concentrations of 0%, 0.5%, 0.75%, 1.0%, 1.5%, 2.0%, 2.5%, 3.0%, 3.5%, 4.0%, 4.5%, 5.0%, 5.5%, 6.0%, 6.5%, 7.0%, 7.5%, 8.0%, 8.5%, and 9.0% was prepared. Three replications of each treatment were used. After inoculation with seed cultures the initial growth was detected by measuring optical density at 600nm. Then the treatments were incubated for 48 hours at 35°C.
3. Results and Discussion
Microbial growth is directly dependent on how temperature affects cellular enzymes. With increasing temperatures, enzyme activity increases until the three-dimensional configuration of these molecules is lost because of denaturation of their protein structure. On the other hand, as the temperature is lowered toward the freezing point, enzyme inactivation occurs and cellular metabolism gradually diminishes. At 0°C temperature biochemical reactions cease in most cells. For normal growth and maintenance the temperature at a range of 25-30°C are usually used for incubation of the great majority of Streptomyces, Nocardia, and Micromonospora. Frequently somewhat different temperatures are used for seed production and for antibiotic formation under factory conditions. Pathogenic organisms require 37°C, and thermophiles need 50-60°C or even higher temperatures [39]. Figure 1 represents the different growth patterns of two actinomycetes isolates viz., JUBM-35-NS-1 (Streptomyces sp.) and JUBM-35-NS-2 (Nocardia sp.). The temperature range for growth was found from 25°C to 45°C with respective OD600 values of 0.06 and 0.03 for isolate JUBM-35-NS-1 and it was 0.1 and 0.115 for isolate JUBM-35-NS-2. The maximum growth was observed at 30°C and 35°C respectively for the isolates JUBM-35-NS-1 and JUBM-35-NS-2. At 4°C and 50°C temperatures growth of both isolates was found to be ceased completely. At 30°C temperature the isolate JUBM-35-NS-1 showed better growth with an OD600 value of 0.07 while at temperature 35°C the isolate JUBM-35-N-2 showed better growth with an OD600 value of 0.26. Thus the strain JUBM-35-NS-2 is more vigorous at its optimum temperature than the strain JUBM-35-NS-1. Waksman [37] reported a number of species isolated from soil with their temperature optimum for growth varied from 25°C to 55°C, the maximum for most species being at 40°C and the minimum below 18-20°C. Another study by Haines [11] on measurements of growth rate of Actinomyces from both soil and cold stores showed that the majority of the strains had their optimum temperature at 37°C, capable of growth from 40 to 50°C. One strain, however, had its optimum temperature for growth less sharply defined at 20-30°C. Tawfik and Ramadan [33] studied in Saudi Arabia and reported that the optimum temperature for growth and antibiotic production by two strains of Streptomyces was 30°C.
Growth and survival of bacteria are also greatly influenced by the pH of the environment. The specific range for bacteria is between 4 and 9, with the optimum being 6.5 to 7.5. Different bacteria prefer different pH range. Streptomyces are neutrophilic showing better growth in between pH 6.0 and 8.0 [18]. The pH range for growth of the isolate JUBM-35-NS-1 was detected from 5.5 with OD600 of 0.04 to 7.5 with OD600 of 0.055 and it was found ranging from pH 5.5 with OD600 of 0.075 to pH 8.0 with OD600 of 0.085 in case of isolate JUBM-35-NS-2. In case of isolate JUBM-35-NS-1 the best growth was recorded at pH 6.0 and 6.5 (OD value 0.065) and in case of JUBM-35-NS-2 the best growth was recorded at pH 6.5 (OD value 0.14). The optimum growth was observed in between pH 6.0 to 7.5 with OD600 range of 0.065 - 0.055 for isolate JUBM-35-NS-1, and it was in between pH 6.5 to 8.0 with OD600 range of 0.14 - 0.13. Thus it has been evident that the isolate JUBM-35-NS-2 is more vigorous with wide ranging pH adaptability (Figure 2). Farmer [8] reported that Nocardia favours an alkaline pH 8.0 for their growth. Ten species of Streptomyces from Finland have been reported to show maximum growth at a pH range of 4.0-5.5 [20].
James et al. [17] reported an optimum temperature of 45°C and that of pH in between 5.5 to 6.5 for highest growth rate of Streptomyces thermoviolacus. Kim et al. [19] reported a new species of Streptomyces growing abundant at a pH range of 4.3-7.3 and between a temperature of 25 and 37°C but not at 45°C. An antifungal Streptomyces has been found to show its optimum activity at 30°C and pH 6.5 [15]. Palanichamy et al. [25] has reported Streptomyces species from Chennai coastal region with optimum temperature of 28-30°C and pH of 7.6-8.0. Bhavana et al. [4] have reported that optimum mycelial growth and antibiotic production by Streptomyces carpaticus from the Vishakhapatnam sea coast of Bay of Bengal occurred at pH 7.2 and at 30°C. Higher production of bioactive metabolites by Streptomyces species SRDP-TK-07 isolated from Talakaveri, Western Ghats of Karnataka, India was observed at temperature 45°C, initial pH 7.0 and salinity of 2% [28].
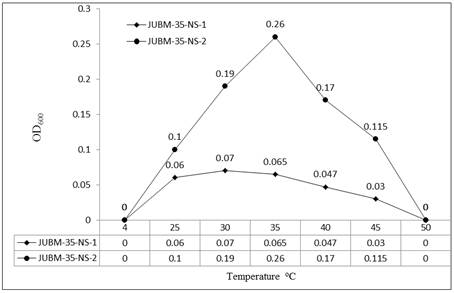
Figure 1. Effect of temperature on the growth of Actinomycetes isolates JUBM-35-NS-1 and JUBM-35-NS-2.
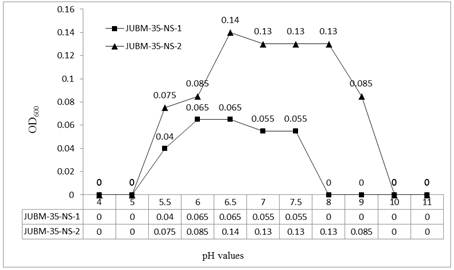
Figure 2. Effect of pH on Actinomycetes isolates JUBM-35-NS-1 and JUBM-35-NS-2.
The isolates were also evaluated for their ability to withstand salt stress by growing at NaCl concentrations of 0%, 0.25%, 0.5%, 0.75%, 1.0%, 1.5%, 2.0%, 2.5%, 3.0%, 3.5%, 4.0%, 4.5%, 5.0%, 5.5%, 6.0%, 6.5%, 7.0%, 7.5%, 8.0%, 8.5% and 9.0%. The best growth (OD600 value 0.095) showed by isolate JUBM-35-NS-1 was in absence of NaCl and it was observed in case of JUBM-35-NS-2 at 0.75% NaCl with OD600 value of 0.2 (Figure 3). Growth arrest was recorded at NaCl concentration of 8.5% and 2.5% respectively for the isolate JUBM-35-NS-1 and JUBM-35-NS-2. Thus the NaCl tolerance limit for growth of JUBM-35-NS-1 was up to 8% (OD value 0.0325) and that for the isolate JUBM-35-NS-2 was 2% (OD value 0.03). The growth of isolate NS-1 dropped from OD600 0.095 at 0% NaCl to OD600 value of 0.055 with increased NaCl concentration to 0.5%. Then it continued till the NaCl concentration of 8.0% without further declination. It testifies the isolate’s capacity to withstand salinity up to that level and thus this is a halotolerant strain. However, in case of isolate JUBM-35-NS-2 NaCl at lower concentration exerted a stimulatory effect on its growth up to 0.75% NaCl and thereafter growth of the isolate declined sharply to complete cessation at NaCl concentration of 2.5%. Hence, the isolate JUBM-35-NS-2 is not a halotolerant strain though its maximum growth is dependent on low salt concentration (Figure 3). Tresner et al. [34] reported that Streptomyces strains can tolerate NaCl concentration from 4% up to 13%. They also reported that in evaluating the relationships of NaCl tolerance to various taxonomic features, higher tolerance was statistically associated with the "yellow" and possibly the "white"-spored Streptomyces. The isolate of this present study is in agreement with Tresner et al. [34]. Moreover, Salle [30] noted that minute amounts of metallic ions or salts stimulate growth of microorganisms whereas concentrations produce an inhibitory effect. In addition to affecting osmotic pressure, high salt concentrations tend to denature proteins and obligate halophiles possess specialized enzymes that are in their active configuration only at high salt concentrations [2].
Streptomyces spp. VITSVK9 showed the maximum growth with highest biomass at 30°C and at 5% of NaCl concentration in the medium and at pH 7.0; and the growth of the strain was inhibited in the absence of NaCl in the medium [31].
Moreover these halophilic microorganisms can be conveniently grouped according to their NaCl requirements for growth [35]. Extreme halophiles have been shown to grow in saturated NaCl concentration and fail to grow below 12% NaCl. Larsen [22] has defined as moderate halophiles are organisms growing optimally between 5 to 20% NaCl concentrations.
Ripa et al. [29] recovered a new Streptomyces strain in 2009 from Bangladesh soil with its activity maxima at temperature range of 25-45°C and at NaCl concentration range of 0.5-3.0% at alkaline pH.
A strain of Streptomyces grancidicus with optimum activity at 40°C, pH 7.0 and 1.5% NaCl has been reported in 2015 from Indian soil [21]. Effect of different physico-chemical parameters included pH, temperature, incubation period and NaCl concentration on the growth of a Streptomyces isolate from India showed growth optima at 45°C, pH 7.0 with failure of growing beyond 6.0% NaCl concentration [27]. Three species of Streptomyces from Malaysia have been found to show optimum growth at temperatures of 25, 30 and 37°C at pH 7.0 with most preferred NaCl concentration of 3.0% by all three isolates [12].
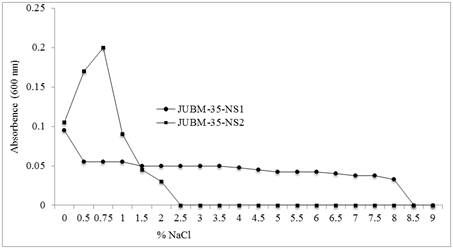
Figure 3. Effect of NaCl on the growth of Streptomyces sp. (JUBM-35-NS-1) and Nocardia sp. (JUBM-35-NS-2).
Nocardia strains reported from South Africa showed growth at pH range of 7.0-10.0; at 30 and 37°C, but not at pH 4.3 and 45°C; and in the presence of 4% NaCl; some strains grow weakly in the presence of 7% NaCl [7]. The investigation on optimal growth temperature and NaCl tolerance of a Nocardiopsis alba strain isolated from bioaerosol of a mushroom compost facility revealed growth optimum at temperature range of 28-37°C and NaCl tolerance up to 10% without any growth at temperature below 25°C and above 45°C [26].
4. Conclusion
Population of microorganisms during composting indicates the performance and quality of the compost. Reports on microbial community during composting reveal wide variations caused by the feedstock differences and the vast variety of composting conditions (i.e., type of raw material, facility design, aeration rate, pH, C/N ratio, temperature and moisture content). Therefore, monitoring of the microbial succession during composting process may provide important information of compost quality [3].
Proper composting promotes the development of a number of saprophytic soil microorganisms. Among others, species of Streptomyces and Nocardia have been reported by several authors in various parts of the world from compost and straw samples [1], [6], [9], [14], [21], [23] [24], [32]. In the present study optimum levels of culture conditions were determined for Streptomyces and Nocardia isolates which have been evident playing effective role in degrading rice straw and compost development for mushroom cultivation (data not shown here).
References
- R. Ashraf, F. Shahid and T. A. Ali, "Association of fungi, bacteria and actinomycetes with different composts," Pak. J. Bot., vol. 39, no. 6, pp. 2141-2151, 2007.
- R. M. Atlas and R. Bartha, "Microbial Ecology: Fundamentals and Applications," Addison–Wesley Publishing Company Inc., London, 1981.
- N. A. A. Bakar, N. Ibrahim, M. H. A. Rahman and N. A. Shakri, "Microbial population assessment during IMO-composting production," Malaysian J. Microbiol., vol. 11, no. 1, pp. 47-53, 2015.
- M. Bhavana, V. S. S. L. P. Talluri, K. S. Kumar and S. V. Rajagopal, "Optimization of culture conditions of Streptomyces carpaticus (mtcc-11062) for the production of antimicrobial compound," Int. J. Pharm. Sci., vol. 6, no. 8, pp. 281-285, 2014.
- V. Dhananjeyan, N. Selvan and K. Dhanapal, "Isolation, characterization, screening and antibiotic sensitivity of actinomycetes from locally (Near MCAS) collected soil samples," J. Biol. Sci,, vol. 10, pp. 514-519, 2010.
- M. F. Eida, T. Nagaoka, J. Wasaki and K. Kouno, "Isolation and characterization of cellulose-decomposing bacteria inhabiting sawdust and coffee residue composts," Microbes Environ., vol. 27, no. 3, pp. 226–233, 2012.
- G. J. Everest, A. E. Cook, M. le Roes-Hill and P. R. Meyers, "Nocardia rhamnosiphila sp. nov., isolated from soil," Syst. Appl. Microbiol., vol. 34, pp. 508-512, 2011.
- R. Farmer, "A selective medium for Nocardia," Proc. Oklahoma Acad. Sci., pp. 285-287, 1961.
- H. W. Feng, Y. E. Zhi, W. W. Shi, L. Mao and P. Zhou, "Isolation, identification and characterization of a straw degrading Streptomyces griseorubens JSD-1," Afr. J. Microbiol. Res., vol. 7, no. 22, pp. 2730-2735, 2013.
- M. Goodfellow and S. T. Williams, "Ecology of actinomycetes," Ann. Rev. Microbiol., vol. 37, pp. 189-216, 1983.
- R. B. Haines, "The influence of temperature on the rate of growth of saprophyticActinomyces," J. Exp. Biol., vol 9, pp. 45-60, 1932.
- A.A. Hamid, S. Ariffin, S. A. S. Mohamad, "Identification and optimal growth conditions of actinomycetes isolated from mangrove environment," Malaysian J. Analytical Sci, vol. 19, no. 4, pp. 904-910, 2015.
- J. G. Holt, "Bergey's Manual of Determinative Bacteriology," 9th Edition, Williams & Wilkins, Baltimore. ISBN 0-683-00603-7, 1994.
- H. Insam,andM. de Bertoldi, "Microbiology of the composting process," In: Compost Science and Technology, L. F. Diaz, M. de Bertoldi, W. Bidlingmaier and E. Stentiford (eds), Waste Management Series, Volume 8, Elsevier Ltd., ISBN: 978-0-08-043960-0, pp. 25-48, 2007.
- M. R. Islam, Y. T. Jeong, Y. J. Ryu, C. H. Song and Y. S. Lee, "Isolation, identification and optimal culture conditions of Streptomyces albidoflavus C247 producing antifungal agents against Rhizoctonia solani AG2-2," Microbiology, vol. 37, no. 2, pp. 114-120, 2009.
- M. N. Jahan,G. M. Khairuzzaman, F. T. Zohora, M. A. Hossain, R. Y. Shilpi and M. A. Akond, ‘‘Isolation of actinomycetes strains from straw and compost samples of Savar, Dhaka, Bangladesh,"Jahangirnagar University J. Biol. Sci., vol. 2, no. 1, pp. 57-64,2013.
- P. D. A. James, C. Edwards and M. Dawson, "The effects of temperature, pH and growth rate on secondary metabolism in Streptomyces thermoviolaceus grown in a chemostat," J. Gen. Microbiol., vol. 137, pp. 1715-1720,1991.
- I. S. Kim, H. T. Kim, A. C. Ward, M. Goodfellow, Y. C. Hah, and K. J. Lee, "Numerical identification of a Streptomyces strain producing thiol protease inhibitor," J Microbiol Biotecbnol, vol. 2, pp. 220-225, 1992.
- S. B. Kim, C. N. Seong, S. J. Jeon, K. S. Bae and M. Goodfellow, "Taxonomic study of neutrotolerant acidophilic actinomycetes isolated from soil and description of Streptomyces yeochonensis sp. nov.," Int. J. System. Evol. Microbiol. vol. 54, pp. 211-214, 2004.
- M. Kontro, U. Lignell and A. Nevaleinen, "pH effects on 10 Streptomyces spp. growth and sporulation depend on nutrients, Lett. App. Microbiol., vol. 41, pp. 32-38, 2005.
- A. Krishnan, and S. S. Kumar, "Optimization of alpha amylase from marine actinomycetes- Streptomyces granidicus_ASD-KT852565," Int. Res. J. Pharm., vol. 6, no. 10, pp. 729-735, 2015.
- H. Larsen, "Halophilic and halotolerant microorganisms: an overview and historical perspective," FEMS Microbiol. Rev., vol. 39, pp. 3-7, 1986.
- A. L. M. Low, S. A. S. Mohamad and M. F. F. Abdullah, "Taxonomic diversity and antimicrobial activities of actinomycetes from manure composts," Res. J. Microbiol., vol. 10, no. 11, pp. 513-522, 2015.
- S. Mohammed, and D. Muleta, "Microbial dynamics during composting of coffee waste (husk) in jimma, southwestern Ethiopia," World J. Pharm. Res., vol. 4, no. 6, pp. 206-227, 2015.
- V. Palanichamy, A. Hundet, B. Mitra and N. Reddy, "Optimization of cultivation parameters for growth and pigment production by Streptomyces spp. isolated from marine sediment and rhizosphere soil," Int. J. Plant Animal Environ. Sci., vol. 1, no. 3, pp. 158-170, 2011.
- M. Pasciak, M., K. Pawlik, A. Gamian, B. Szponar, J. Skora, B. Gutarowska, "An airborne actinobacteria Nocardiopsis alba isolated from bioaerosol of a mushroom compost facility," Aerobiologia, vol. 30, pp. 413–422, 2014.
- P. Prasad, T. Singh, S. Bedi, "Characterization of the cellulolytic enzyme produced by Streptomyces griseorubens (Accession No. AB184139) isolated from Indian soil," J. King Saud Univ. – Sci., vol. 25, pp. 245–250, 2013.
- K. N. Rakesh, N. Dileep, S. Junaid and T. R. P. Kekuda, "Optimization of culture conditions for production of antibacterial metabolite by bioactive Streptomyces species srdp-tk-07," PHARMANEST, vol. 5, no. 1, pp. 1809-1816, 2014.
- F. A. Ripa, F. Nikkon, S. Zaman and P. Khondkar, "Optimal conditions for antimicrobial metabolites production from a new Streptomyces sp. RUPA-08PR isolated from Bangladeshi Soil, Mycobiology, vol. 37, no. 3, pp. 211-214, 2009.
- A. J. Salle, "Fundamental Principles of Bacteriology," 6th Edition, TATA McGraw-Hill Publishing Co. Ltd., Bombay, New Delhi, 1967.
- K. Saurav and K. Kannabiran, "Diversity and Optimization of process parameters for the growth of Streptomyces VITSVK9 spp. isolated from Bay of Bengal, India," J. Natural Environ. Sci., vol. 1, no. 2, pp. 56-65, 2010.
- P. F. Strom, "Identification of thermophilic bacteria in solid-waste composting," Appl. Environ. Microbiol., vol. 50, no. 4, pp. 906-913, 1985.
- K. A. Tawfik and E. M. Ramadan, "Factors affecting the biological activity of Streptomyces aureofaciens MY18 and Str. roseviolaceus MR13," J. King Abdul Aziz Univ.: Sci., vol. 3, no. 5-19, 1991.
- H. D. Tresner, J. A. Hayes, and E. J. Backus, "Differential tolerance of Streptomycetes to sodium chloride as a taxonomic aid, Appl. Microbiol., vol. 16, no. 8, pp. 1134-1136, 1968.
- A. Ventosa, J. J. Nieto and A. Oren, "Biology of moderately halophilic aerobic bacteria," Microbiol. Mol. Biol. Rev., vol. 62, pp. 504-544, 1998.
- J. Viji and P. Neelanarayanan, "Efficacy of lignocellulolytic fungi on the biodegradation of paddy straw," Int. J. Environ. Res., vol. 9, no. 1, pp. 225-232, 2015.
- S. A. Waksman, "Cultural studies of species of Actinomyces," Soil. Sci.. vol. 8, pp. 71–215, 1919.
- S. A. Waksman, "The Actinomycetes; Vol. 1. Nature, occurrence and activities" The Williams &WilkinsCompany, Baltimore, 1959.
- S. A. Waksman and H. A. LeChevalier, "Guide to the classification and identification of the actinomycetes and their antibiotics," The Williams & Wilkins Company, Baltimore, 1953.



