
Public Health and Preventive Medicine, Vol. 1, No. 2, June 2015 Publish Date: May 28, 2015 Pages: 66-72
Biochemical Criteria of Naegleria Species Isolated from the Egyptian Aquatic Environment
Wafaa M. Hikal1, 2, *, Ahmad Z. Al-Herrawy2, Mahmoud M. Bahgat3, Abd-Elhafez H. Mohammed4, Ameen A. Ashour4
1Department of Biology, Faculty of Science, University of Tabuk, Tabuk, Saudi Arabia
2Parasitology Lab., Water Pollution Research Department, National Research Centre, Dokki, Giza, Egypt
3Therapeutic Chemistry Department, National Research Centre, Dokki, Giza, Egypt
4Zoology Department, Faculty of Science, Ain Shams University, Cairo, Egypt
Abstract
The free-living amoebae Naegleria species (spp.) have been recognized as etiologic agents of amoebic encephalitis, keratitis, otitis, lung lesions and other skin infections mainly in immuno-compromised individuals. In this study, morphological and biochemical characterization of Naegleia strains isolated from the Egyptian aquatic environment were surveyed. Some Naegleria species were cultivated on non-nutrient agar. Isolated strains of Naegleria were identified based on the morphology of trophic and cyst forms in addition to temperature and flagellation test. Biochemical characterization of the isolated amoeba strains using quantitative and qualitative (SDS-PAGE) assays as well as qualitative determination of proteolytic activity in zymograph analysis. Potentially pathogenic free-living amoebae were isolated from all of the examined water sources. Colorimetric assays showed protease activity in heat-tolerant isolates of Naegleria. All pathogenic isolates exhibited higher protease activity than non-pathogenic ones did. The zymographic protease assays showed various banding patterns for different strains of Naegleria.
Keywords
Free-Living Amoebae, Naegleria, Flagellation Test, Proteases, Drinking Water Sources
Received: April 8, 2015
Accepted: April 19, 2015
Published online: May 27, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
Contents
1. Introduction 2. Materials and Methods 2.1. Samples and Sampling Sites 2.2. Isolation and Morphologic Identification of Naegleria Spp. from Water Samples 2.3. Flagellation Test 2.4. Biochemical Characterization of Isolated Naegleria spp. 2.5. Statistical Analysis 3. Results 3.1. Morphological Characterization of Genus Naegleria 3.2. Prevalence of Genus Naegleria in Different Water Sources 3.3. Quantitatively Absolute Enzyme Activity in NaegleriaIsolates 3.4. QualitativelySilver Stain Profile of Resolved Naegleria Protein 3.5. Qualitatively Proteolytic Activity in Lysates of Different Naegleria Isolates Visualized by Gelatin SDS-PAGE 4. Discussion 4.1. Prevalence of Heat Tolerant Free-Living Amoebae in Different Types of Water 4.2. Biochemical Characterization of Isolated Free-Living Amoebae 5. Conclusion
1. Introduction
Infection of the central nervous system by free-living amoebae is an unusual event. Free-living amoebae are presented worldwide in fresh water as well as in marine water. Moreover, they have been recovered from various domestic water systems such as drinking tap water (Michel et al., 1998), cooling towers (Barbaree et al., 1986), swimming pools (Rivera et al., 1983), hydrotherapy baths (Scaglia et al., 1983) and hospital water networks (Thomas et al., 2006). Waterborne transmission, acquired through forceful inhalation of surface waters or poorly maintained swimming pools, is uncommon (Karanis et al., 2007). The genus Naegleria includes more than 30 species of freelivingamoebae that are extensively distributed in soil and freshwaterenvironments (CDC, 2010; Laseke et al, 2010). Naegleria fowleri is the only species of thisgenus known to be pathogenic to humans (Visvesvara et al, 2007). The disease most associated with N. fowleri infection is primary amoebic meningo-encephalitis (PAM), an acute, severe, necrotizing, and hemorrhagic form of meningo-encephalitis.PAM infection requires that N. fowleri trophozoites or cysts enter the central nervous system. Infection begins when contaminated water directs through the nose, usually through swimming or diving. Inside the nasal passage, travel amoebae reach the olfactory mucosa, along the olfactory nerve fibers, and through the perforated plate in the brain.Inside the brain, N. fowleri amoebae feed on red blood cells, white blood cells, and brain tissue. PAM is almost always fatal, usually killing its victims within 3–7 days after the onset of symptoms (CDC, 2010; Cabanes et al, 2001; John, 1982). Pathogenic FLA are not dependent upon a host for transmission and spread, nor does host-to-host transmission of these amoebic diseases occur. They feed by phagocytosis, mainly on bacteria, fungi and algae (Bass and Bischoff, 2001). They have the ability to multiply and grow well in tropical climate and in water body with high temperatures of 40-45ºC (De Jonckheere, 2006).There are few data on the occurrence of these pathogenic free-living amoebae in the aquatic environment of Egypt. So, the main objective of the present work is to illustrate the occurrence and identification of pathogenic free-living amoebae Naegleria spp in different types of water using morphological characteristics. A secondary objective is to characterize the potential pathogenicity of the isolated strains using biochemical assays.
Table 1. Samples and sampling sites.
| Locality | Water type |
| Cairo | Nile and tap |
| Giza | Nile and tap |
| Qalubeya | Nile and tap |
| Behera | Nile and tap |
| Gharbeya | Nile and tap |
| Dakahleya | Nile and tap |
| Helwan | Nile and tap |
| Kafr-Elshikh | Tap |
| Sharkeya | Tap |
| Minofeya | Tap |
Figure 1. Governorates of the Nile Delta, Egypt.
2. Materials and Methods
2.1. Samples and Sampling Sites
Water samples (2 liters each) were collected from different localities in Delta region, Egypt (Figure 1) for the detection and isolation of freshwater amoebae(Table 1). Samples were collected from the Nile Riverand tap water in clean, dry autoclavable polypropylene containers and sent to the laboratory in icebox and processed at the same day of collection.
2.2. Isolation and Morphologic Identification of Naegleria Spp. from Water Samples
Collected water samples (1 liter each) were concentrated by using the membrane filtration technique. One liter of each water sample was filtered through a nitrocellulose membrane filters (0.45 μm pore size and 47 mm in diameter) (Whatman, WCN type, Cat No. 7141-104) (Gradus et al., 1989). After filtration the membranes were separately inverted face to face on the surface of a non-nutrient (NN) agar plates previously seeded with 100 μl Escherichia coli suspension. All the inoculated plates were incubated at 40°C for one week with daily microscopic examination for the presence of any amoebic growth (Hikal, 2015). Identification of the obtained Naegleriaspp. were achieved according to the morphological characteristics of both trophic and cyst stages (Pussard and Pons, 1977, Hikal, 2010, Al Herrawy et al., 2013).
2.3. Flagellation Test
The obtained amoebic trophozoites were gently scraped from the surface of agar plates with a bacteriological loop and suspended in a test tube containing 5 ml distilled water and incubated at 37°C for 30 minutes. Every 10 minutes one drop from the content of the tube was suspended in the concavity of a clean glass hanging drop slide and examined under the microscope for the formation of temporary flagella (Behets et al., 2003).
2.4. Biochemical Characterization of Isolated Naegleria spp.
Grown amoebae were harvested from cultured NN agar plates by scraping of the agar surface in eppendorf tubes containing 0.5 ml sterile Page's amoebae saline. The harvested amoebae were centrifuged at 1500 rpm for 10 min. The supernatant was discarded and the final pellet was re-suspended in 100 μl Page's amoebae saline and homogenized for 5 minutes in a tissue grinder. After that, the homogenate was transferred to a fresh eppendorf tube and centrifuged at 14000 rpm for 10 min. The supernatant was aspirated, divided into aliquots and stored at -80ºC till being used. These steps were repeated for each sample (Khan et al. 2000).
2.4.1. Quantitative Assays for Proteinase Activity Using Chromomeric Substrates
An aliquot was taken from samples prepared and stored at -80°C as mentioned above. The protease activities in different Naegleriawere quantitatively measured using the trypsin-like proteases specific substrate (Boc-Val-Leu-Gly-Arg-PNA L-1195, Bachem Bioche-mica, Heidelberg, Germany) at λmax 405 nm using Sun Rise reader (TECAN, Austria) according to (Iwanaga, 1994; Bahgat and Ruppel, 2002; Bahgat et al., 2006). The intensity of the yellow colour was directly proportional to the enzyme activity.
2.4.2. Qualitative Determination of Proteolytic Activity inZymograph Analysis (Gelatin Sodium-Dodecyl Sulphate Polyacrylamide Gel Electrophoresis, SDS-PAGE Gels)
The proteolytic activity of Naegleriaisolates were characterized by zymography on SDS-polyacrylamide gels copolymerized with gelatin (Bahgat et al., 2006).
2.5. Statistical Analysis
All obtained data were analyzed by the student's t-test using the Graph Pad InStat Soft ware.
3. Results
3.1. Morphological Characterization of Genus Naegleria
Genus Naegleria represented the amoebo-flagellates whose members could transform from amoebae to flagellate forms. Differentiation of Naegleria from other amoebae was based on their characteristic eruptive movement of the amoebic form, and their ability to transform to flagellates.
3.2. Prevalence of Genus Naegleria in Different Water Sources
Heat tolerance and morphological features were used in differentiating members of genus Naegleria from other amoebae. Heat-tolerant Naegleria species were isolated from only 24.6and 16.7% of the examined Nile water and Tap water, respectively (Figure 2).
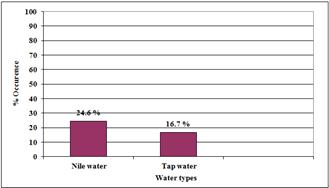
Figure 2. Occurrence of Naegleria at different water sources.
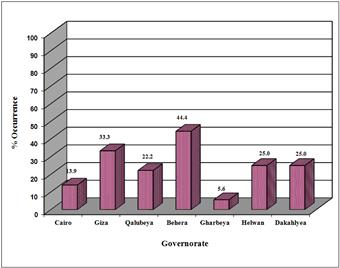
Figure 3. Occurrence of Naegleria in Nile water samples from different sites.
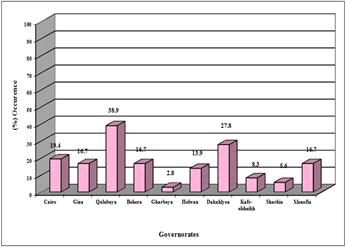
Figure 4. Occurrence of Naegleria in tap water samples from different sites.
Nile water samples collected from Behera Governorate showed the highest incidence of heat-tolerant Naegleria species (44.4%), followed by Giza in a percentage of 33.3%. The incidence of heat-tolerant Naegleria species was the same (25.0%) in raw Nile water collected from both Helwan and Dakahleya Governorates. The least incidence of Naegleria species in the Nile water samples occurred in Gharbeya (5.6%) and Cairo (13.9%) Governorates (Figure 3).
The highest incidence of heat-tolerant Naegleria species was recorded in tap water samples collected from Qalubeya Governorate (38.9%), followed by Helwan and Cairo in percentages of 27.8 and 19.4 %, respectively. The incidence of heat-tolerant Naegleria species was the same (16.7%) in tap water collected from Giza, Behera and Minofeya Governorates. The lowest incidence of heat-tolerant Naegleria species in tap water from Gharbeya, Sharkeya and Kafr-Elshikh Governorates in percentages of 2.8, 5.6 and 8.3%, respectively (Figure 4).
3.3. Quantitatively Absolute Enzyme Activity in NaegleriaIsolates
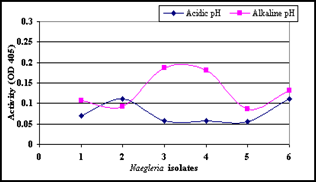
Figure 5. Tryptase activity in individual Naegleria isolates at both acidic and alkaline pH values.
3.4. QualitativelySilver Stain Profile of Resolved Naegleria Protein
The proteolytic profile of bacterial control sample without Naegleria was completely different from that of Naegleria isolates. The bacterial control sample without Naegleria had strong bands at molecular weights 124, 103, 90, 74, 66, 60, 53, 49, 45, 41, 35, 32, 27, 24, 21 and 15 kDa.The proteolytic profiles of prepared lysates from both Naegleria isolates number 1 and 3 (isolated from tap water) had 2 common bands at molecular weights 121 and 89 kDa. The lysate of isolate number 1 showed nine strong bands at 135, 121, 89, 69, 60, 50, 23, 19 and 15 KDa and three weak bands at 45, 35 and 27 kDa, while the lysate of isolate number 3 was resolved into seven strong bands at 143, 121, 109, 89, 75, 63, 48 kDa and two weak bands at 24 and 20 kDa. Naegleria isolates number 2 and 4 (isolated from tap and Nile water) showed different proteolytic activities. The lysate of Naegleria isolate number 2 showed four strong bands at 141, 112, 60, and 50 kDa, while Naegleria isolate number 4 showed five strong bands at 139, 121, 91, 61, 52 KDa and one weak band at 44kDa. Naegleriaisolates number 5 and 6 (isolated from Nile water) had a common band at 44 kDa. The lysate of Naegleria isolate number 5 showed bands at molecular weights 139, 119, 107, 61, 51, 44, 32 and 14 kDa, while in Naegleria isolate number 6 the lysate was resolved into five strong bands at 141, 121, 89, 73, 60 and 49 kDa and two weak bands at 105 and 44 kDa (Figure 6).
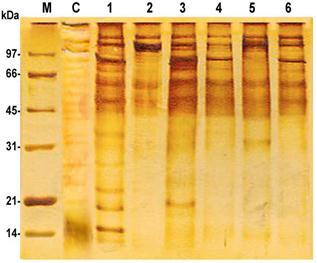
Figure 6. Protein profile of Naegleria isolates followed by silver stain, whereC: negative control bacteria; (1, 2, 3, 4, 5 and 6): N. spp.; M: marker.
3.5. Qualitatively Proteolytic Activity in Lysates of Different Naegleria Isolates Visualized by Gelatin SDS-PAGE
The proteolytic profile of prepared lysates from bacterial control samplecontaining no Naegleria was totally different from that of different Naegleria isolates. Lysates from bacterial control sample had molecular weights at 147, 110, 87, 70, 59, 53, 46, 31 and 23 kDa.The proteolytic activities of prepared lysates of Naegleria isolates showed activity at both alkaline and acidic pH values. Isolates number 1 and 3 (isolated from tap water) showed different profiles. Naegleria isolate number 1 visualized proteolytic activity bands at 143, 115, 96, 78, 59, 51, 40 and 31 kDa. In Naegleria isolate number 3, the proteolytic activity visualized bands at 164, 133, 112, 90, 73 and 63 kDa. Naegleria isolates number 2 and 4 (isolated from tap and Nile water) also showed different proteolytic profiles. In Naegleria isolate number 2 the gelatin digestion bands were evidenced at 137, 113, 92, 76, 64, 58, 42 and 30 kDa.Naegleria isolate number 4, the proteolytic activity visualized bands at 167, 141, 117, 89, 69, 57 and 52 kDa. Naegleria isolate number 5 (isolated from Nile water) showed three strong proteolytic bands at 139, 113 and 93 kDa and four weak bands at 74, 62, 54 and 48 kDa (Figure 7).
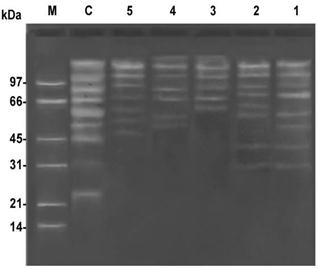
Figure 7. Serine-like protease activity in Naegleria isolates was visualized in gelatin SDS-PAGE,whereC: negative control bacteria; (1, 2, 3, 4, and 5): N. spp.; M: marker.
4. Discussion
The present study deals with the natural distribution of pathogenic Naegleria in the aquatic environment at different localities in the Nile Delta region, Egypt. Of special interest were the isolates capable of proliferating at temperatures of 37°C and above, as well as the biochemical characterization of these organisms. Previous studies on this subject in Egypt are non-existent. Therefore this study represents the first field investigation initiated in Egypt to estimate the presence of potentially pathogenic Naegleria in the Nileand tap waters by using culture and biochemical techniques.
4.1. Prevalence of Heat Tolerant Free-Living Amoebae in Different Types of Water
In the present study the incidence of Naegleria reached 24.6% in the examined Nile water samples. Other workers in Egypt also detected Naegleria in 66.7 and 60% of the examined freshwater samples by Hilali et al. (1994) and Hamadto et al. (1993), respectively. In USA, other workers detected Naegleria from pond water and James River in percentages of 56 and 46%, respectively (John and Howard, 1995; Ettinger et al., 2003). Also in Saudi Arabia, Al-Herrawy and Al-Rashied (1995) detected Naegleria in 47.22% of the examined freshwater samples. In Thailand, Nacapunchai et al. (2001) recorded an incidence (28.6%) lower than that recorded in our results.In the present work, the incidence of Naegleria reached 16.7% in the examined tap-water samples. Other workers in Egypt detected Naegleria in 9.7% of the examined tap water (Hilali et al., 1994).
4.2. Biochemical Characterization of Isolated Free-Living Amoebae
Proteases are enzymes that catalyze the hydrolysis of peptide bonds in a broad spectrum of important biological reactions including the pathogenesis of parasitic disease (McKerrowet al., 1993). In the present work, Naegleria isolates showed different proteolytic activity bands ranging from 30 to 167 kDa. Other workers showed that five out of fifteen enzymes: aldehyde oxidase (ALDOX), aldolase (ALD), a-glycerophosphate dehydrogenase (a-GPDH), xanthine dehydrogenase (XDH), and glutamate oxaloacetate transaminase (GOT), were undetectable in the pathogenic strains, while the other enzymes: esterase (EST), fumerase (FUM), glucose-6-phosphate dehydrogenase (G-6-PDH), glucose phosphate isomerase (GPI), isocitate dehydrogenase (IDH), lactate dehydrogenase (LDH), leucine aminopeptidase (LAP), malic enzyme (ME), glucose phosphomutase (GPM), and malate dehydrogenase (MDH), were detected (Tiewcharoen et al., 2004). Other workers in Malaysia (Amin, 2004) showed that N. fowleri possesses two high molecular weight proteases on gelatin gels at molecular weights 128 and 170 kDa.
5. Conclusion
The incidence and prevalence of the pathogenic free-living amoebae in different populations using morphological and biochemical diagnostic tools will provide baseline data against which the risk factors associated with waterborne transmission can be identified. The isolated species of Naegleriacould provoke variable degrees of infections to the swimmers. The culture method is cheaper and easier than biochemical techniques that are faster for the detection of free-living amoebae.
References
- Al-Herrawy, A., Bahgat, M., Mohammed, A., Ashour, A. andW. Hikal (2013). Morpho-Physiological and Biochemical Criteria of Acanthamoeba spp. Isolated from the Egyptian Aquatic Environment. Iranian J Parasitol., 8 (2): 302-312.
- Al-Herrawy, A. Z. and K. A. S. Al-Rasheid (1995). Isolation of thermo-tolerant Naegleria from a freshwater course in Riyadh (Saudi Arbia). J. Vet. Sci., 1: 79-83.
- Amin, M. N. (2004). Proteinases in Naegleria Fowleri (strain NF3), a pathogenic amoeba: a preliminary study.Trop. Biomed., 21:57-60.
- Bahgat, M. and A.Ruppel(2002). Biochemical comparison of the serine protease (elastase) activities in cercarial secretions from Trichobilharzia ocellata and Schistosoma mansoni. Parasitol. Res., 88: 495-500.
- Bahgat, M., Sorgho, H. and J. B. Ouédraogo(2006). Enzyme-linked immunosorbent assay with worm vomit and cercarial secretions of Schistosoma mansoni to detect infections in an endemic focus of Burkina Faso.J. Helminthol., 80:19-23.
- Barbaree, J. M., Fields, B. S., Feeley, J. C., Gorman, G. W. and W. T. Martin (1986). Isolation of protozoa from water associated with a legionellosis outbreak and demonstration of intracellular multiplication of Legionella pneumophila. Appl. Environ. Microbiol., 51: 422-424.
- Bass, P. and P. J. Bischoff (2001). Seasonal variability in abundance and diversity of soil gymnamoebae a short transect in southeastern USA. J. Eukaryot. Microbiol., 48: 475-479.
- Behets, J., Seghi, F., Declerck, P., Verelst, L., Duvivier, L., Van Damme, A. and F. Ollevier (2003). Detection of Naegleria spp. and Naegleria fowleri: a comparison of flagellation tests, ELISA and PCR. Wat. Sci. Technol., 47: 117-122.
- Cabanes, P., Wallet, F., Pringuez, E. and P. Pernin(2001). Assessing the Risk of Primary Amoebic Meningoencephalitis from Swimming in the Presence of Environmental Naegleria fowleri. Applied & Envir. Microbiol., 67:7:2927.
- CDC (Centers for Disease Control and Prevention), 2010.www.dpd.cdc.gov/dpdx/HTML/FreeLivingAmebic.htm(accessed Oct. 31, 2011).
- De Jonckheere, J. F. (2006). Molecular identification of free-living amoebae of the Vahlkampfiidae and Acanthanoebidae isolated in Arizona (USA). Eur. J. Protistol., 43: 9–15.
- Ettinger, M. R., Webb, S. R., Harris, S. A., McIninch, S. P., Garman, G. C. and B. L. Brown (2003). Distribution of free-living amoebae in James River, Virginia, USA. Parasitol. Res., 89: 6-15.
- Gradus, M. S., Koenig, S. B., Hyndiuk, R. A. and J. De Carlo (1989). Filter-culture technique using amoebae saline transport medium for the noninvasive diagnosis of Acanthamoeba keratitis. Am. J. Clin. Pathol., 92: 682-685.
- Hamadto, H. H., Aufy, S. M., El-Hayawan, I. A., Saleh, M. H. I. M. and Nagaty (1993). Study of free-living amoebae in Egypt. J. Egypt. Soci. Parasitol., 23: 631-637.
- Hikal, W. M. (2010): Biochemical and molecular characterization of pathogenic free-living amoeba in the aquatic environment. ph. D. thesis, fac. Sci., Ain Shams Univ., Egypt.
- Hikal, W. M. (2015). Detection of Acanthamoeba Species from Water Tanks in Saudi Arabia. Asian Academic Research J. Multidisciplnary, (1): 42-49.
- Hilali, M., Ashmawy, K., Samaha, H., Draz, A. A., Abu El-Wafa, S. A. and Salem, A. (1994). Preliminary studies on amoebic pathogens isolated from water and sewage with respect to Naegleria and Acanthamoeba. J. Egypt. Vet. Med. Ass., 53: 215-224.
- Iwanaga, Y. (1994). Studies on host-parasite relationship between the Puerto Rican strain of Schistosoma mansoni and Biomphalaria snails. Southeast Asian J. Trop. Med. Public Health, 25: 509-15.
- John, D. T. (1982). Primary amoebic meningo-encephalitis and the biology of Naegleria fowleri. Ann. Rev. Microbiol., 36: 101-123.
- John, D. T. and M. J. Howard (1995). Seasonal distribution of pathogenic free-living amoebae in Oklahoma water. Parasitol. Res., 81: 193-201.
- Karanis, P., Kourenti, C. and H. Smith(2007). Waterborne transmission of protozoan parasites: A worldwide review of outbreaks and lessons learnt. J. Wat. Health, 5: 1-38.
- Khan, N. A., Jarroll, E. L., Panjwani, N., Cao, Z. and T. A. Pager (2000). Proteases as markers of differentiation of pathogenic and non-pathogenic Acanthamoeba. J. Clin. Microbiol., 38: 2858-2861.
- Laseke, I., Korte, J., Lamendella, R., Kaneshiro, E.S., Marciano-Cabral, F. and D.B. Oerther ( 2010). Identification of Naegleria fowleri in Warm Ground Water. Aquifers. Jour. Envir. Quality,39:147.
- Mckerrow, J. H., Sun, E., Rosenthal, P. J. and J. Bouvier(1993). The proteases and pathogenicity of parasitic protozoa. Annu. Rev. Microbiol., 47: 821-853.
- Michel, R., Muller, K. D., Amann, R. and E. N. Schmid (1998). Legionella-like slender rods multiplying within a strain of Acanthamoeba spp. isolated from drinking water. Parasitol. Res., 84: 84-88.
- Nacapunchai, D., Kino, H., Ruangsitticha, C., Sriwichai, P., Ishih, A. and M. Terada(2001). A brief survey of free-living amoebae in Thailand and Hamamatsu district, Japan. Southeast Asian j. Trop, Med. Public Health, 32: 179-82.
- Pussard, M. and R. Pons (1977). Morphologie de la paroi kystique et taxonomie du genre Acanthamoeba (Protozoa, Amoebida). Protistol., TXIII: 557-598.
- Rivera, F., Ramirez, P., Vilaclara, G., Robles, E. and F.Medina(1983). A survey of pathogenic and free-living amoebae inhabiting swimming pool water in Mexico City. Environ. Res., 32: 205-211.
- Scaglia, M., Strosselli, M., Grazioli, V., Gatti, S.; Bernuzzi, A.M. and J.F. De Jonckheere (1983). Isolation and identification of pathogenic Naegleria australiensis (Amoebida, Vahlkampfidae) from spa in northern Italy. Appl. Environ. Microbiol., 46: 1282-1285.
- Thomas, V., Herrera-Rimann, K., Blanc, D. S. G. and Greub (2006). Biodiversity of amoebae and amoebae-resisting bacteria in a hospital water network. Appl. Environ. Microbiol.,72: 2428-2438.
- Tiewcharoen, S., Komalamisra, N. and V. Junnu (2004). Zymogram patterns of Naegleria spp. isolated from natural water sources in taling chain district, Bangkok. South East Asian j. Trop. Med. Public. Health, 35: 275-280.
- Visvesvara, G. S., Moura, H. andF. L. Schuster (2007). Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri and Sappinia diploidea. FEMS Immunol. Med. Microbiol., 50: 1–26.



