
American Journal of Food Science and Health, Vol. 1, No. 3, August 2015 Publish Date: Jul. 16, 2015 Pages: 82-85
The HOXD Locus in the Pathogenesis of Celiac Disease
Procino Alfredo*
Department of Neurosciences, Reproductive and Odontostomatologic Sciences, Medical School Federico II, Naples, Italy
Abstract
Celiac disease (CD) is immune-mediated enteropathy that occurs in genetically predisposed subjects following ingestion of an alcohol-soluble component of gluten, gliadin, found in some cereals (wheat, barley, rye, spelled, etc.). HOX, are 39 transcription factors mostly involved in the regulation of embryonic development and regulates the cell memory program through the interaction with non coding RNA. I have already titled this network as "The Rosetta stone" of human biology; therefore, does HOX network able to control change in the cell memory program during CD?
Keywords
Celiac Disease, Homeobox, lncRNA, HOTAIR
Received:April 4, 2015
Accepted: June 25, 2015
Published online: July 15, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
Contents
1. Introduction
Celiac disease (CD) is a multigenic disease characterized by derangement of adaptive and innate immune responses to wheat gliadins [1]. Some gliadin peptides, deamidated by tissue transglutaminase (i.e., A-gliadinP57-68) bind to HLA DQ2/DQ8 molecules and induce an adaptive Th1 pro-inflammatory response. Other gliadin peptides (i.e., P31-43) are able tostart both stress and innate immune response [2].
2. Main Body
Damage to the intestinal CD mucosa is mediated by inflammation due to both adaptive and innate immune responses (IL-15 is a major mediator of the innate immune response) and by proliferation of crypt enterocytes, that causes crypt hyperplasia and mucosal remodelling, hallmarks of CD. One hypothesis consider some gliadin peptides acting as viruses on the intestinal mucosa by entering the endocytic cell compartment and interfering with the correct maturation of endocytic vesicles [3]. Together these results point to direct damaging effects of undigestedgliadin peptides on intestinal epithelial cells. The effects of these peptides are extremely disruptive to intestinal mucosa and it has not been elucidated. Maybe, in CD, a constitutive genetic/epigenetic alteration represent a predisposing condition, to the damaging effects of gliadin undigested peptides and viral infections like [2]. Recently, two genomic regions were identified associated with CD [4] and located on chromosome 2: ITG4/UBE2E3 (2q31.3) and CTLA4/ICOS/CD28 (2q33.2). The genes present in this area, are in physical contiguity with the HOXD locus present on chromosome 2q31-3210. Class I homeobox genes (HOX), are 39 transcription factors, mainly involved in the regulation of embryonic development program and arranging of the human body plan; The HOX genes are characterized by a sequence of 183 nucleotides encoding a homeodomain of 61 amino acid that binds to DNA, biological gripper (Fig. 1), activating or repressing specific genes [5, 6]. Moreover, They are organized into four chromosomal clusters or loci (HOXAChr7p15.3, HOXBChr17q21.3, HOXCChr12q13.3 and HOXDChr2q31), each having 9-11 genes. Considering their position into the locus and similarity of homeobox sequence, corresponding genes, of the four clusters, can be aligned with each other in 13 paralogous groups (Figure 2) [7]. The HOX network is active in adult human tissues and organs, controls the spatial-temporal generation ofbiological structure expected during embryonic development andregulates the cell memory program (Figure 2).Genes related to the cell memory program, finely control the cell phenotype.Epigenetic change in the cell memory program is critical in the onset and evolution cancer [8]. Three genes families control this process:Polycomb (H3K27m3), able to block the interaction DNA-chromatin leading to silencing of HOX genes. Trithorax genes (H3K4m3), able to induce the mRNA transcription through an open configuration of DNA-chromatin interaction and leads to activation of HOX genes.Finally, the HOX genes involved in the control of the phenotype through a specific gene program, mainly by means the accurate regulation of the mRNA transcription (Fig. 3) [9].In the last decade, it has been identified 231 lncRNA into the network; in particular, the lncRNAHOX antisense intergenic RNA (HOTAIR) was identified in 2007. HOTAIR is transcribed from HOXC locus located at chromosome 12q13.13, in a position intergenic and antisense to the flanking HOXC11 and HOXC12 genes [10, 11, 12, 13]. HOTAIR is the first lncRNA that control the gene expression in trans. In details, it is able to blockthe transcriptional control of the HOXD locus on chromosome 2, including the unique Polycomb Responsive Element (PRE) to dateidentified in the genome. Recently, 30 genes were identified into the small bowel able to control the epithelial cell differentiation and celiac disease pathogenesis [14]. Several studies suggest that HOTAIR acts regulating epigenetic states by recruiting chromatin-modifying complexes to specific target sequences that ultimately leads to suppression of different genes [15] and also HOTAIR is a powerful predictor of poor prognosis in colon cancer, mostly controlling the EMT processes [16].
3. Conclusion
In conclusion, based on the literature and the results obtained, I have hypothesized that HOTAIR and HOX genes would induce an alteration of the epithelial cell memory program, consequent to the ingestion of the gluten through the food, maybe due to a deregulation of the EMT process.
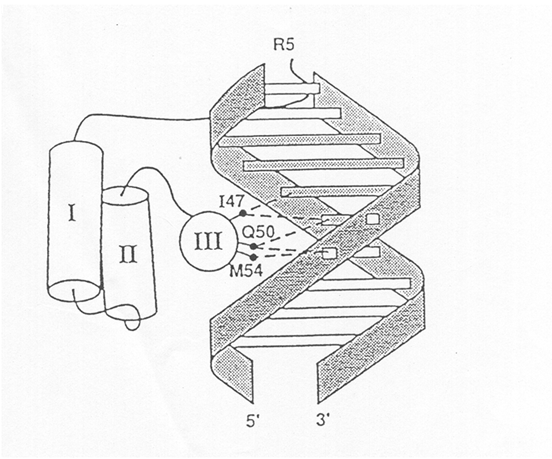
Fig. 1. Alfa-helix homeodomain structure. The homeodomain is able to bind DNA with its secondary structure at triple-helical. The third helix, recognizes a specific nucleotide sequence on the DNA and binds it. Therefore, the role of the homeodomain is to promote or repress the gene expression located to downstream of the sequence identified.
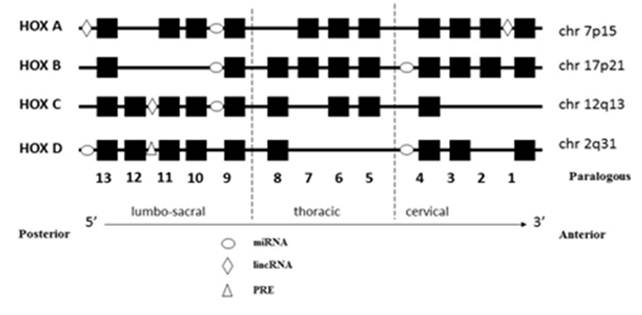
Fig. 2. Schematic representation of the HOX gene network. The Hox genes are organized into four chromosomal clusters or loci: HOX A at 7p15.3, HOX B at 17q21.3, HOX C at 12q13 and HOX D at 2q31. HOX and genes are 39 transcriptional regulators of embryonic development.
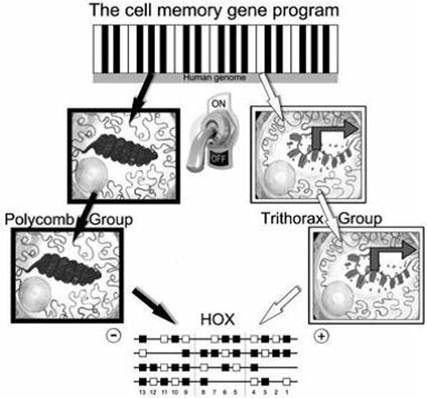
Fig. 3. Three genes family regulates the cell memory program: (i) Trithorax genes, able to induce mRNA transcription through (ii) HOX genes activation and (iii) Polycomb genes, able to block mRNA transcription through HOX genes silencing.
References
- Abadie V, Sollid ML, Barreiro LB and Jabri B. Integration of Genetic and Immunological Insights into a Model of Celiac Disease Pathogenesis Annu. Rev. Immunol. 2011. 29:493– 525.
- Iacomino G, Fierro O, D'Auria S, Picariello G, Ferranti P, Liguori C, Addeo F, Mamone G. Structural analysis and Caco-2 cell permeability of the celiac-toxic A-gliadin peptide 31- 55. J Agric Food Chem. 2013 Feb 6;61(5):1088-96.
- Troncone R, Auricchio S. Rotavirus and celiac disease: clues to the pathogenesis and perspectives on prevention. J PediatrGastroenterolNutr. 2007;44:527-8.
- Dubois PC, Trynka G, Franke L, Hunt KA, Romanos J, et al. 2010. Multiple common variants for celiac disease influencing immune gene expression. Nat. Genet. 42:295–302.
- Ferber S, Halkin A, Cohen H, Ber I, Einav Y, et al. (2000) Pancreatic and duodenal homeobox gene 1 induces expression of insulin genes in liver and ameliorates streptozotocin-induced hyperglycemia. Nat Med 6: 568-572.
- Foucher I, Volovitch M, Frain M, Kim JJ, SouberbielleJC, et al. (2002) Hoxa5 overexpression correlates with IGFBP1upregulation and postnatal dwarfism: evidence for an interaction between Hoxa5 and Forkhead box transcription factors. Development 129: 4065-4074.
- Barnes TM, Kohara Y, Coulson A, Hekimi S (1995) Meiotic recombination, noncoding DNA and genomic organization in Caenorhabditiselegans. Genetics 141: 159-179.
- Bantignies F, Cavalli G (2006) Cellular memory and dynamic regulation of polycomb group proteins. CurrOpin Cell Biol 18: 275-283.
- GehringW.J., Hiromi Y (1986) Homeotic genes and the homeobox. Annu Rev Genet 20: 147-173.
- Functional demarcation of active and silent chromatin domains in human hox loci by noncoding RNAs. 2007. Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Cell 129, 1311–1323.
- Long noncoding rnahotair regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. 2011. Kogo, R.; Shimamura, T.; Mimori, K.; Kawahara, K.; Imoto, S.; Sudo, T.; Tanaka, F.; Shibata, K.; Suzuki, A.; Komune, S. Cancer Res. 71, 6320–6326.
- Long non-coding RNAs in cancer progression. Tano, K.; Akimitsu, N. 2012 Front. Genet.3, 219.
- The sequence, structure and evolutionary features of hotairin mammals. He, S.; Liu, S.; Zhu, H. 2011 BMC Evolut. Biol.11, 102.
- Gluten affects epithelial differentiation-associated genes in small intestinal mucosa of coeliac patients. Juuti-Uusitalo K, Maki M, Kainulainen H, Isola J, Kaukinen K 2007 ClinExpImmunol 150: 294–305.
- The lincRNAHotair is required for epithelial-to-mesenchymal transition and stemness maintenance of cancer cell lines. Pádua Alves C1, Fonseca AS, Muys BR, de Barros E Lima Bueno R, Bürger MC, de Souza JE, Valente V, Zago MA, Silva WA Jr. Stem Cells. 2013 Dec;31(12):2827-32.
- Long non-coding RNA HOTAIR is a powerfu predictor of metastasis and poor prognosis and is associated with epithelial-mesenchymal transition in colon cancer.Wu ZH, Wang XL, Tang HM, Jiang T, Chen J, Lu S, QiuGQ, PengZH, Yan DW.Oncol Rep. 2014 Jul;32(1):395-402.



