
International Journal of Plant Science and Ecology, Vol. 1, No. 4, August 2015 Publish Date: Jul. 9, 2015 Pages: 162-166
The Content of Nonenzymic Antioxidants in Different Organs Alhagi pseudalhagi Grown Under Conditions of Chronic γ – Radiation
E. S. Jafarov1, *, G. A. Gojayeva1, A. K. Jafarli1, H. G. Babayev2
1Radiobiology laboratory,Institute of Radiation Problems, Azerbaijan National Academy of Sciences, Baku, Azerbaijan
2Biochemical laboratory, Institute of Botany, Azerbaijan National Academy of Sciences, Baku, Azerbaijan
Abstract
In this paper were investigated features of accumulation and changes in the content of anthocyanins, flavonoids and carotenoids in different organs Alhagi pseudalhagi (Bieb.), grown in conditions of chronic γ-radiation. An attempt was made to clarify the role of these nonenzymic antioxidants in conditions of radiation stress. On the basis of a slight increase in pigment content was concluded that the protective responses in the plant are involved nonenzymic antioxidants less effective.
Keywords
Alhagi pseudalhagi, Chronic γ- Radiation, Antioxidants, Flavonoids, Anthocyanins, Carotenoids
Received:June 17, 2015
Accepted: June 19, 2015
Published online: July 7, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
1. Introduction
It is known that the action of various stressors leads to decrease in biodiversity wild plants and productivity of agricultural plants. Therefore, the study of intracellular molecular mechanisms allowing plants to adapt to unfavorable factors is one of the actual problems plant physiology. It is believed that as a result of multiple metabolic changes in the stress factor action formed a so-called adaptive response and the plant originally resistant to the effects of [5,7].
It should be noted that among the factors responsible for the adaptation of plants to adverse environmental conditions, the composition and properties of the pigments is not the last. To date had the idea that the different pigments, as quenchers of reactive oxygen species, act as antioxidants [1,2]. This increase in antioxidant levels in adverse environmental conditions can be mediated by the accumulation of reactive oxygen species. Therefore, an increase of their contents can be regarded as a defensive response of plants under such conditions. It can be assumed that the difference in the rate of biosynthesis of antioxidants in ontogeny determines their content.
It should be noted that plants have sufficient resistance to oxidative damage. This manifests itself when exposed to various external factors, or the sudden change in the physiological state of the plant. Plant resistance is due, primarily, by the presence in their cage highly protective systems, which consists of high-enzymatic components and low molecular weight antioxidants.
Various studies conducted in the field investigation show that the effective functioning of the antioxidant systems under stress is determined by many factors. Such factors particularly include the types of plants, the degree, duration, and character of stress, the age and conditions of plant growth, as well as the state of antioxidant system and the initial level of antioxidant activity [6,15].
The results of studies on the protective reactions of plants under the influence of stress factors show their diversity. In some cases, protection of plant participate low molecular weight metabolites such as glutathione, ascorbic acid, anthocyanins, flavonoids, carotenoids, proline, tocopherol, ubiquinone etc. [8]. In individual cases high-molecular antioxidant enzymes such as superoxide dismutase, catalase, glutathione reductase, various peroxidases and other enzymes cycle Halliwell - Asada can make a significant contribution to the protection of plants against stress [4].
The aim of the present work is to study the characteristics of accumulation and changes in the content of nonenzymic antioxidants such as anthocyanins, flavonoids, carotenoids in different organs Alhagi pseudalhagi (Bieb.), growing in conditions of chronic γ-radiation.
2. Material and Methods
To solve this problem have been chosen as the control and the experimental plots. At the time of the study, the mean exposure dose in the control and experimental plots was (10.5 ± 1.2) μR/hr and (550 ± 47) μR/hr, respectively. Soil and climatic conditions of the control and experimental plots were almost identical.
Determination of qualitative and quantitative content of radionuclides in soil samples showed that the main radionuclides, polluting experimental plots are 40K, 232Th and 226Ra. Specific activities of these radionuclides were respectively 8240, 4500 and 49000 Bq / kg [13].
Alhagi pseudalhagi is a perennial plant. We used a biennial sample of this plant. Samples of different organs for fixation and extraction of pigments were taken at budding phase of plants. Total pigments were determined in the freshly harvested plant organs.
Anthocyanin and carotenoid concentration was determined by the method of Sims, Gamon [14]. For this, samples of the individual plants were homogenized in 80% acetone solution. After three days of exposure extract was filtered and prepared a special blend. Special mixture of 1 ml contains 800 mkl 80% acetone, 180 mkl Tris buffer (pH 7,8) and 20 mkl plant extract. Standard solution with a volume of 1 ml contained 800 mkl 80% acetone, 200 mkl 1M Tris buffer (pH 7,8).
Further using a spectrophotometer (Ultrospec 3300 pro Amersham Bio-sciences) were determined optical density. Optical density of anthocyanins were determined at wavelengths 537 nm, 647 nm and 663 nm, and the optical density of сarotenoids - 470 nm, 537 nm, 647 nm and 663 nm. On this basis, with using equations were determine the concentration of these pigments.
For quantitative determination of total flavonoid content in different organs of the plant raw material sample were ground to a particle size passing through a sieve with openings of 1 mm diameter. Chopped raw material was transferred to a flask with 70% ethanol, and the flask were attached a reflux condenser and heated in a boiling water bath for 60 min [11].
The extract were filtered after cooling. The extraction was repeated twice under the same conditions with 70% ethyl alcohol for 60 minutes at the second contact phase and 30 minutes at the third (Solution A). Solution A was transferred to a volumetric flask. Were poured a 2% solution of aluminum chloride in 95% ethanol, 1 drop of 5% acetic acid and the solution volume was adjusted to 95% ethyl alcohol to the mark (solution B). After 40 minutes on a spectrophotometer at a wavelength of 414 nm in a cuvette with a layer thickness of 10 mm, the optical density of the solution AB It has been determined. For the preparation of the reference solution A solution was transferred to a volumetric flask, was poured 1 drop of 5% acetic acid and the solution volume was adjusted to 95% ethyl alcohol until the mark [11].
In parallel, the optical density of the solution of routine has been determined.
Total anthocyanins (its concentration in μmol/ml) were determined spectrophotometrically at wavelengths λ = 537 nm, 647 nm and 663 nm according to the formula presented in [14]:
Kant. =0.08173 ·A537 – 0.00697· A647 – 0.002228·A663
(where ![]() and
and ![]() - optical density at wavelengths of 537, 647 and 663 nm, respectively).
- optical density at wavelengths of 537, 647 and 663 nm, respectively).
The concentration of carotenoids was determined based on the content of anthocyanins and chlorophylls a, b according to the formula [14]:
Kkarot.={ A470 - [ 17.1· (Kxl.a+Kxl.b) - 9.479·Kant.]}/119.26
(where ![]() and
and ![]() the concentration of anthocyanins, chlorophylls a, b in μmol/ml, respectively).
the concentration of anthocyanins, chlorophylls a, b in μmol/ml, respectively).
Wherein the concentrations of chlorophylls a and b (in μmol/ml) were determined by the formulas [14]:
Kxl.a = 0.01373 A663 – 0.000897 A537 – 0.003046 A647
Kxl.b = 0.02405 A647 – 0.004305 A537 – 0.005507 A663
For determining the total number flavonoids in plant samples were used a method developed S.S. Lambayeva [11]. Using differential spectrophotometry at a wavelength λ = 414 nm the total flavonoids conrent was determined in percentages to rediscount at rutin and absolutely dry raw material by the formula:
![]()
![]()
(where ![]() - optical density of the test solution,
- optical density of the test solution, ![]() - optical density of the solution GSO rutin,
- optical density of the solution GSO rutin,![]() - weight in grams of the starting material,
- weight in grams of the starting material, ![]() - weight in grams GSO rutin,
- weight in grams GSO rutin, ![]() - the dilution coefficient of the test solution (
- the dilution coefficient of the test solution (![]() =1250),
=1250), ![]() - the dilution coefficient of the GSO rutin solution (
- the dilution coefficient of the GSO rutin solution (![]() 2500) ,
2500) , ![]() - weight loss in percent on drying of the starting material).
- weight loss in percent on drying of the starting material).
Experiments were carried out in double biological and triple analytical replicates, which gave results with an error of 0 to ± 20%. The figures show the mean values of the measured values. Statistical processing was performed by standard methods of variation statistics. The significance of differences of control and experimental results was assessed using Student’s t – distribution of [9]. The differences were significant at |t| ˃2 (p<0.05).
3. Discussion of Results
As is known, practical human activity has led to what in some sites of the world were formed locally contaminated areas with different radionuclides. Formation of such sites on the territory of our Republic is associated with the extraction and processing of oil and gas. The area with high background radiation, we have chosen as experimental plot to study the effects of ionizing radiation on wild plants growing on it.
Observations of chronically irradiated populations of wild plants have shown that in an area with high background radiation grows best Alhagi pseudalhagi (Bieb).
Given that the state of plant populations at contaminated sites primarily evaluated their morphological features, we carried out systematic observations of chronically irradiated A.pseudalhagi. These observations showed that both in the biometric indicators and in the vegetation period of the experimental plants as compared with control samples there are some differences [3]
Note that the use of physiological and biochemical characteristics of assimilating organs for early diagnosis of the condition of plants in ecosystems acceptable. Since these bodies largely reflect the processes of development and reproduction of plants. Furthermore, they are the most sensitive to various environmental changes. However, from the perspective of the internal potential and its mobilization to adverse conditions, attempts are based on early diagnosis will not be sufficient.
It is known that for protection against the effects of adverse environmental factors plants have effective defense systems, which consists of different antioxidants [12]. Therefore, the study of different antioxidants which play a key role in protecting plants against stress factors, as well as the study of regularities accumulation and changes their content in different plant organs as a result of plant responses to the stressor, today is a priority.
Based on these considerations, we have attempted to clarify the features of accumulation and interdependent functioning of some low molecular weight antioxidants (flavonoids, anthocyanins and carotenoids) in different organs of A. pseudalhagi under conditions of chronic γ - radiation. Note that today almost no publications to clarify the contribution of different groups of low molecular weight compounds in the functioning of the cell antioxidant defense system.
Our data for the determination of flavonoids in different organs of the plant and change them in conditions of chronic γ-radiation are shown in Fig. 1.
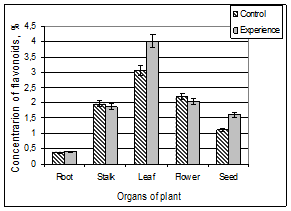
Figure 1. The content of flavonoids in different organs of A. pseudalhagi.
From the diagrams, mainly attracts attention the presence of flavonoids in all plant organs and their uneven distribution in different organs. In general, the content of total flavonoids in the aerial parts of the plants was significantly higher than in the roots. Content of pigments in the leaves, flowers, stems, seeds and roots of control plant relative to the dry weight were 3.05 ± 0.18%, 2.21 ± 0.43%, 1.97 ± 0.13%, 1.13 ± 0.05% and 0.38 ± 0.06%, respectively. As can be seen, the leaves of this plant are characterized by the highest (30.5 ± 1.8 mg / g) and the roots of the lowest (3.8 ± 0.6 mg / g) flavonoid composition.
As regards participation of radiation in the process, it should be noted that ionizing radiation did not lead to a significant change in the content of flavonoids in the roots, stems and flowers of the plant. Exception is leaves and seeds. In conditions of chronic γ – radiation the content of flavonoids in these organs increased approximately 31 and 42%, respectively.
It is interesting that, if do not take into account the slight variations, the nature of the distribution of flavonoids in different organs of the experimental and control samples are almost identical. In other words, in this case, as in the case of the control plant sample the roots have the least amount flavonoids, in the stems their content is increased and becomes the maximal in the leaves. Further, going over to the flowers, their content is reduced and in the seed becomes even smaller.
The next step of our research was to investigate the distribution of anthocyanins in different organs A. pseudalhagi.
Note that a specialized plant metabolism characterized by extreme plasticity. According to modern concepts, this is due to the fact that during the metabolism in plants takes place chemical modification. Among the components of specialized metabolism phenol compounds are among the most numerous. Most of higher plants especially metabolic accumulation of anthocyanins hardly been studied. Therefore, the study of species - specific and organ - specific features of the accumulation of flavonoids, as well as the regulation of metabolism of these compounds is of great scientific and practical interest.
It is appropriate to note that the results obtained different researchers with respect to the content of anthocyanins in individual plant organs are ambiguous. Usually the qualitative composition of anthocyanins for the given plant is specific and relatively is stable. The composition of pigments may vary depending on the kind of plant and growing conditions [10].
Analysis of the results obtained by us for A. pseudalhagi shows that the flowers of this plant have a maximum number of anthocyanin: (9.11 ± 1.47) μmol / ml. In this case, the roots of the plants have a minimal amount of anthocyanins: (1.79 ± 0.52) μmol / ml. The amounts of this pigment in the stems were within the limits (3.39 ± 0.73) μmol / ml (Figure 2).
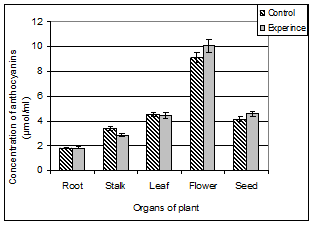
Figure 2. The content of anthocyanins in different organs of A. pseudalhagi.
At first glance, for plants grown in conditions of chronic radiation exposure, features of the anthocyanins distribution by different organs and their accumulation in specific organ does not differ from that of the control plants. However, the analysis of results show that ionizing radiation causes an insignificant inhibition of anthocyanins synthesis in the stems. At the time occurs a slight stimulation of anthocyanins synthesis in the flowers. Let us recall that the ionizing radiation both in the leaves and in the seeds of the plant resulted in the accumulation of antioxidant flavonoids also.
Ionizing radiation did not cause significant changes in the content of anthocyanins in the roots, leaves and seeds of this plant.
Another interesting fact is that between the accumulation capacity of anthocyanins and flavonoids, there is a positive correlation. The difference is that if flowers exhibit the highest activity to the process of anthocyanins synthesis, that the leaves of the plant exhibit the highest activity to the flavonoids synthesis.
Cognition biological significance of the secondary synthesis of substances such as carotenoids and rational use of carotenoid-containing plants is impossible without a detailed study of the laws of accumulation, specificity and variability of the composition and content of these substances. Taking this into account, we have attempted to determine the content of carotenoids and to evaluate the role of ionizing radiation during their accumulation in different organs of A. pseudalhagi.
Data obtained by us on the content of carotenoids are shown in Figure 3.
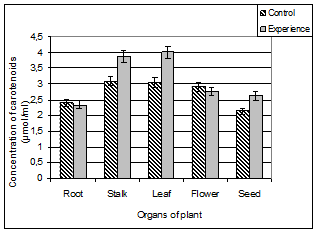
Figure 3. The content of carotenoids in different organs of A. pseudalhagi.
As can be seen from the diagrams the plant bodies of control and test samples are characterized by different distributions of carotenoids. In this case, the control sample of plants in the stems, leaves and flowers has almost the same amount of carotenoids: (3.09 ± 0.57) μmol/ml, (3.05 ± 0.45) μmol / ml and (2.91 ± 0.49) μmol / ml, respectively. The roots and seeds of the plants contain a relatively low amount ((2.41 ± 0.53) μmol / ml and (2.13 ± 0.32) μmol / ml, respectively) of yellow pigments.
Distribution of carotenoids in organs of the test plants differs from control. Stem, leaves and seeds of the test variant has a higher stock of this pigment. Since the carotenoid composition in these organs were respectively 1.2, 1.3 and 1.25 times more than the control plant organs. Understandably, carotenoids, playing the role of antioxidants can be increased in conditions of stress. Then the change of their content can be taken as an indicator of the physiological state of the plant.
Summarizing the results, we can conclude that the different bodies react differently to chronic exposure of γ-radiation. In this the ionizing radiation practically does not cause accumulation of low molecular weight antioxidants in plant roots and its seeds in condition of high background radiation the content of all investigated antioxidants significantly are increased.
Apparently, the plant for preserving the viability in unusable environments, to some extent, tries mobilizing his low molecular weight antioxidant internal potential, which in turn is reflected in the increase in the content of antioxidants. Turned out various in chemical nature carotenoids, flavonoids and anthocyanins, despite on localization in the different compartments may play a similar role in the protection of plants from damage.
We assume that a slight increase in the content of flavonoids, anthocyanins and carotenoids in individual organs A. pseudalhagi shows a less effective participation these pigments in a protective response of the plant. In all probability, the relatively high radioresistance of the investigated plants in conditions of chronic γ- radiation due to the effective functioning of antioxidant enzymes than low molecular weight antioxidants.
References
- ChenY., Wei X., Xie H., Deng H. 2008.Antioxidant 2-phenylbenzofurans and coumestan from Lespedeza vigata.J. Natur. Prod., 71(6): 929-932.
- GoulasV., Papoti V.T., Exarchou V.et al.2010.Contribution of flavonoids to the overall radical scavenging activity of olive (Olea europaea L.) leaf polar extracts. Agr. Food Chem. 58(6): 3303 – 3308.
- Jafarov E.S., Orujova J.R., Jafarli A.K. 2005.Study the effect of gamma radiation in small doses on biomorphological, reproductive and resistant feature of Alhagi pseudalhagi and Zygophyllum L. J. Proceedings of National Academy Sciences of Azerbaijan (Biological sciences).3-4: 168-173 .
- Kartashov A. B., Padyukina N.L., Ivanov Yu. B. et al. 2008. The role of antioxidant systems in the adaptation of wild species of plants to salt stress. Plant Physiology. 55: 516-522.
- Kavi Kishor P.B., Sangami S., Amruthai R.N. et al. 2005.Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Current science. 88(3): 424-438.
- Kawanobu S., Wajima T., Zushi K. et al. 2010.Seasonal variations in the maturation period, antocyanin contect and ascorbic acid content in strawberry fruits. Environ. Contr. Biol. 48(4): 175-18.
- Kuznetsov V.V., Shevyakova N.I. 2007.Polyamines and stress tolerance of plants. Plant Stress. Global Science Books. 1(1): 50-71.
- Kuznetsov V.V., Stetsenko L.A., Shevyakova N.I. 2009. Exogenous cadaverine induces oxidative burst and reduces cadaverine conjugate content in the common ice plant. J.Plant Physiol. 166: 40-51.
- Lakin Q. F. Biometrics. M. 1990. Nauka. 352 p.
- Lee D.W. and Gould K.S. 2002. Anthocyanins in leaves and other vegetative organs: an introduction. In: Advances in Botanical Research. Amsterdam: Academic Press. 37: 1-16.
- Lomboyeva S.S., Tankhayeva L. M., Olennikov D.N. 2008. Method for the quantitative determination of total flavonoid content in the ground part Orthilia Secunda L. J. Chemistry of plant raw materials. 2: 65-68.
- Matysik J., Alia Bhalu B., Mohanty P. 2002Molecular Mechanisms of Quenching of Reactive Oxygen Species by Proline under stress in Plants. Curr. Sci.. 82: 525-532.
- Orujova J.R., Jafarov E .S. 2007. Some features of the distribution of natural radio nuclides in different organs of plants growing in an area of high radiation background. J. Radiation Biology and Radioecology.47(2): 241-246.
- Sims D.A., Gamon J.A. 2002. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sensing of Environment. 81(2-3): 337-354.
- Skupien K., Ochmian I., Grajkowski J., Krzywy-Gawronska E. 2011. Nutrients, antioxidants and antioxident activity of organically and convertionally grown raspberries. Appl. Bot. and Food Qual. 84(1): 85-89.



