
International Journal of Plant Science and Ecology, Vol. 1, No. 4, August 2015 Publish Date: Jul. 9, 2015 Pages: 155-161
Variations in Leaf and Fruit Morphological Traits of Sweet Chestnut (Castanea Sativa) in Hyrcanian Forests, Iran
Karimian Atefe, Taheri Abkenar Kambiz*, Torkaman Javad
Department of Forestry, Faculty of Natural Resources, University of Guilan, Rasht, Iran
Abstract
This paper aims to cluster Castanea sativa based on morphological variations of leaves and fruits. Therefore, the main and natural existing sites of sweet chestnut including Shaft, Fouman and Shafaroud were considered in Guilan province. From each site, 10 trees with various morphological traits with at least 100 meters distance from each other were selected. Leaves and fruits were sampled from selected trees. Totally, the number of 30 samples of fruits and leaves were provided individually. The ANOVA results of quantitative traits (e.g. the length, the width of the lamina, special form of the leaf, total and relative length of the petiole, the leaf basal shape, fruit weight, seed weight and healthy seeds weigh) showed significant difference in the three study areas. It was concluded that the morphological traits such as leaf, the width of the lamina, the relative length of the petiole, the leaf tip shape, lamina shape, seed weight and healthy seeds weight were the most important elements in shaping of cluster.
Keywords
Morphological Traits, Leave and Fruit, Castanea Sativa, Principal Component Analysis, Guilan, Iran
Received:April 5, 2015
Accepted: May 21, 2015
Published online: July 7, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
1. Introduction
Natural Forests of northern Iran along with European Remnant Hardwood Forests belong to the Holarctic (TAHERI, 2011) and are categorized in Qurerceto-Fagetea kingdom ((TAHERI & PILEHVAR, 2008). European chestnut with scientific name of Castanea sativa is a species of flowering plant in the family Fagaceae, native to Europe and Asia Minor, and widely cultivated throughout the temperate world(LITTLE, 1953), which is found in the forests of Guilan province specifically in four sites with acidic soil called Veis-roud, Siah-mazgi, Shafa-roud and Ghaleh-roudkhan. In Iran, Castanea sativa was firstly discovered in 1340 by Prof. Hossein Jazireyee. Chestnut is distributed in altitude ranges from 200 to 600 m most likely in west and north aspect with approximately slope of 50 to 70 percent (HEDAYATII et al., 2003). Sadly, the seed collected by villagers for consumption, economic and social problems, including grazing land net type chestnuts minimum levels declined gradually reduced the number of acres have been thus eroding precious treasures as would be expected genetic distance (HEDAYATI, 2003). Certainly, considering more substantial protection and proper management for this species is depended on our knowledge about ecological needs and genetic diversity of this valuable species. One of the oldest classification methods of vegetation is to investigate plants based on morphological characteristics (WANG et al., 2001). Generally, in tree diversity studies, leaf morphology has been targeted as an important subject (BRUSCHI et al., 2003; MC DONALD et al., 2003;KAFFASH et al., 2008; ZARAFSHAR et al., 2009). The interspecies diversity of two species from Prosopis genus situated in the Central and South America was evaluated considering leaf morphological characteristics (<HARRIS et al., 2003). In a research in England focusing on Sorbus genus separation, fruit morphology differences, fruit body structure and the amount of starch in fruit were employed (ALDASORO et al., 1988). Other studies were conducted in relation to the subject in Iran are as follows; Babayee et al. (2010) investigated genetic variation in Zelkova carpinifolia (Pallas) K. Koch, Yousefzade et al. (2010) studied morphological plasticity of Parrotia persica leaves in eastern Hyrcanian forests and Espahbodi et al. 2006) carried out a research on Sorbus torminalis (L) Crantz, using morphological investigation of fruits and leaves. This paper aims to analyze morphological characteristics of chestnut leaves and fruits, in order to determine the most optimum site in Guilan province with consideration of applying appropriate management on preventing the extinction of this rare endangered species.
2. Materials and Methods
At initial point, the three main and natural existing sites of sweet chestnut including Shaft, Fouman and Shafaroud were identified in Gilan province (Figure 1). Site characteristics in the study areas are, illustrated in table 1. In the first half of October in each site, 10 trees with various morphological characteristics with at least 100 meters from each other were selected (MERWIN et al., 1995) and then samples of leaf and fruit were taken from exterior parts of crown among selected trees (AAS et al., 1994). The collected samples of each tree were combined together, the number of 10 fruits and leaves were randomly selected. Totally, 60 samples including 30 fruit and 30 30 leaf samples were gathered for morphological analysis. It should be noted that, the codes from 1 to 10 belong to the sample from Shaft region, 11 to 20 are taken from Fouman area and 21 to 30 are classified in Shafaroud site. The measured traits were including petiole length, lamina length, lamina shape, lamina width, relative petiole length, lamina width of 0.1 leaf length, lamina width of 0.9 leaf length, basal leaf shape, the leaf tip shape, angular leaves base, leaf area index, wall fleshy fruit weigh, large diameter fruit, small diameter fruit, seed number, seed weight, number of healthy seed and weight of healthy seed (ESPAHBODI et al., 2006).
Fig. 1. Location of the study areas.

Fig. 2. Variables measured in the leaves and fruit. LaW: Lamina width, PL:Petiole length, LaL: Lamina length, LeL: Leaf length (U. Serdar and N. Kurt 2011)..
To analyze fruits and leaves data, ANOVA and Duncan test were used. For categorizing the leaves and fruits, the Principal Component Analysis (PCA) and Cluster Analysis were employed. Morphological analyses were performed using the software Pcord.
Table 1. Site characteristics in three study areas.
| site | Elevation (m a.s.l.) | Soil | Geographical location |
| Fouman | 200-350 | Deep and acidic | 37º,5 N- 49º,14 E |
| Shaft | 200-600 | Deep and acidic | 37º, 15 N- 49º,15 E |
| Shafaroud | 250-500 | Deep and acidic | 37º,30 N- 49º, 30 E |
3. Result
The results of ANOVA test showed (table 2) that the morphological traits including lamina length, lamina width, lamina shape, petiole length, relative length petiole, leaf basal shape, leaf area index, fruit weight, seed weight and healthy seed weight were different significantly (p< .01). (Fig2)
Table 2. The average of leaf and fruit morphology traits in study areas.
| Traits Average | Shaft | Fouman | Shafaroud |
| Lamina length | 22.44±1.82b | 22.7±2.257b | 25.04±3.27a |
| Lamina weight | 7.41±0.626b | 6.98±0.795b | 8.66±1.207a |
| Lamina shape | 2.77±0.375a | 3.293±0.515a | 2.892±0.228b |
| Petiole length | 1.85±0.62b | 1.92±0.41a | 1.33±0.176a |
| Relative petiole length | 2.26±0.843a | 2.018±0.899ab | 2.919±0.199b |
| lamina width of 0.1 leaf length | 4.09±0.635a | 3.97±1.155a | 3.84±0.757a |
| lamina width of 0.9 leaf length | 2±0.639a | 1.73±0.731a | 2.17±0.408a |
| basal leaves shape | 0.554±0.075b | 0.565±0.114a | 0.445±0.1a |
| leaf tip shape | 0.27±0.085a | 0.242±0.085a | 0.248±0.023a |
| angular leaves base | 73.8±9.942a | 66.1±13.682a | 69.4±9.442a |
| leaf area index | 54.78±8.017b | 61.495±10.746b | 79.002±20.452a |
| wall fleshy fruit weigh | 22.966±4.99a | 18.64±6.84b | 22.186±5.716b |
| large diameter fruit | 2.56±0.37a | 2.39±0.546a | 2.33±0.47a |
| small diameter fruit | 2.13±0.23a | 3.64±5.42a | 2.5±3.15a |
| seed number per skin | 3a | 3a | 3a |
| seed weight per skin | 10.339±4.745a | 6.52±2.516a | 8.829±3.37b |
| seed number healthy | 2.1±0.737a | 1.6±0.84a | 1.4±0.699a |
| seed weight healthy | 4.7±4.99b | 5.27±2.86a | 70.9±3.5a |
Significant differences among means within a row (α= .01) are marked with different letters

Fig. 3. Sample of leaves and Fruit of chestnut trees.
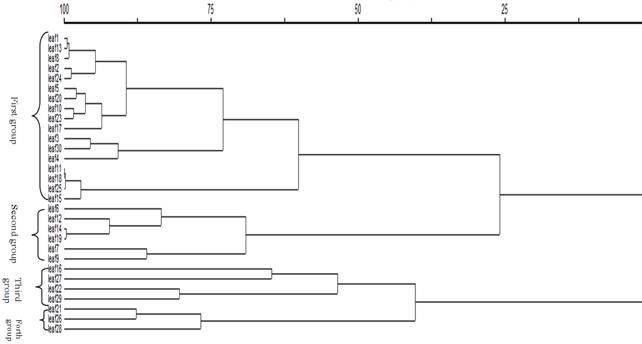
Fig. 4. Dendrogram resulted from cluster analysis of leaves morphological traits.
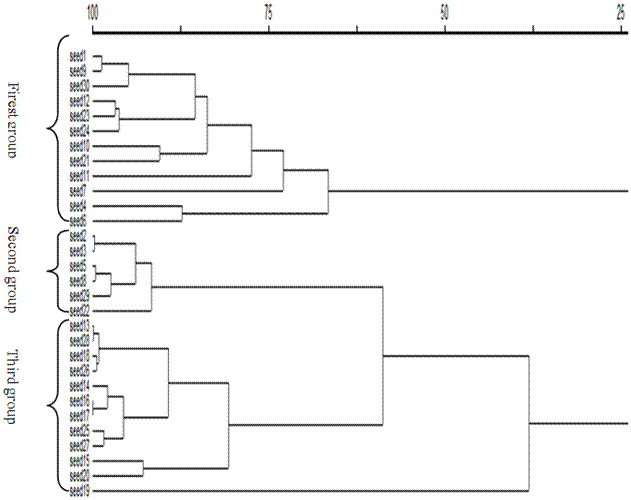
Fig. 5. Dendrogram resulted from cluster analysis of fruits morphological traits.
3.1. Principal Components Analysis of Leaf and Fruit Morphological Characteristics
The PCA was used to determine the similarity and correlation between variables in cluster. The results showed that the first four component of leaf and the first three components of fruit allocated the highest cumulative variance. Generally, the component with the highest cumulative variance is considered as the most important factor in variables grouping (BRUSCHI et al., 2003). The first component was formed by following traits; lamina shape, lamina width of 0.1 leaf length, lamina length, leaf basal shape, leaf angular base, fruit weight, seed weight and healthy seed weight. In relation to preparation the second component, the lamina length, lamina width, lamina width of 0.9 leaf length, leaf area index, fruit largest diameter, seed weight and healthy seed weight were more important than the other morphological characteristics. The lamina shape, relative petiole length, lamina width of 0.9 leaf length, leaf tip shape, fruit weight, fruit smallest diameter, seed weight and healthy seed weight shaped the third component, and finally, the fourth component was constructed by leaf tip shape, lamina length, leaf shape, relative length of petiole and leaf area index. It is obvious that the most important morphological traits which played a significant role in formation of the quad components are the length lamina, lamina shape, leaf tip shape, relative petiole length, lamina shape, lamina width of 0.9 leaf length, leaf area index, healthy seed weight and seed weight.
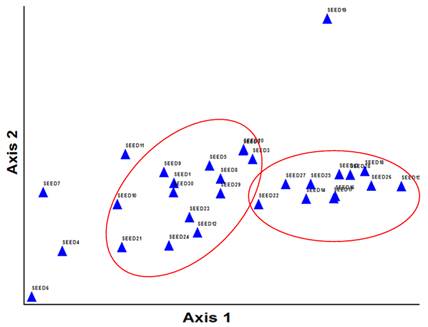
Fig. 6. The scatter plot of fruit resulted from PCA in space coordinate axes based on the first two components.
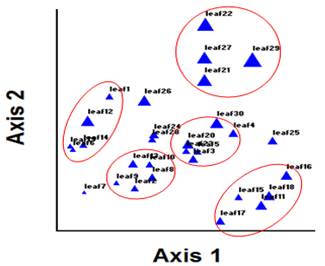
Fig. 7. The scatter plot of leaf resulted from PCA in space coordinate axes based on the first two components.
3.2. Grouping of Leaves and Fruits Based on Morphological Traits
The cluster analysis was performed using the total leaf traits. The results showed that the leaves were categorized into four separate groups. The first group was constituted with encoded leaves of 1, 2, 3, 4, 5 and 8 from Shaft region, the ones with codes of 10, 11, 13, 15, 17, 18 and 20 from Fouman zone and also leaf samples of 23, 24, 25 and 30 from Shafaroud area. The encoded leaves of 6, 7 and 9 from Shaft region and samples with the code of 12, 14 and 19 from Fouman were put into the second group. The third group was shaped by encoded leaves 16 from Fouman and the ones with codes of 22, 27 and 29 from Shafaroud. Finally, the leaves with codes of 21, 26 and 28 from Shafaroud formed the fourth group. It conforms that the leaves in a specific group are morphologically similar to each other. The results of fruit analysis showed that they were categorized into three separate groups (figure 4). The firs group was shaped by encoded fruits of 1, 4, 6, 7, 9 and 10 from Shaft, 11 and 12 from Fouman and 21, 23, 24 and 30 from Shafaroud. The second group was consisted of encoded fruits of 1, 2, 3, 4, 5 and 8 from Shaft region, the ones with codes of 13, 14, 15, 16, 17, 18 and 20 from Fouman zone and also fruit sample of 22 from Shafaroud area. The third group was shaped by encoded fruits of 25, 26, 27, 28 and 29 from Shafaroud and 19 from Fouman (figure 5).
The scatter plots of tree leaves resulted from PCA is showed in figure 7. The figure illustrates that the leaves with various codes are divided into several groups. In fact, leaves with similar morphological traits are classified into a specific group. Also, the scatter plots of the tree fruits are clarified in figure 6. From this figure, it is comprehended that fruits with high resemblance are categorized into a group.
4. Discussion
The leaves and fruits are the most important organ for plant production. The leaf structure provides balance between photosynthesis and transpiration (BRUSCHI et al., 2003). Researches have shown that morphological traits in leaf and fruit have an important role in determining the difference between trees in various habitats (SOKAL & ROHLF, 1995). In this study, among three regions there was significant difference between morphological traits of leaves and fruit, such as lamina length, lamina width, lamina shape, relative petiole length, petiole length, leaf basal shape, leaf area index , fruit weight, seed weight and healthy seed weight. This finding was also confirmed by the results of Serdar and Kurt (2011) and Yusef zadeh et al. (2010).
In addition, the following traits were found notable in shaping of leave group; the lamina shape, the leaf tip shape, relative petiole length, lamina width of 0.9 leaf length, leaf area index and healthy seed weight. It can be concluded that two important parameters; aspect and altitude were mostly effective on the grouping, but slope was not noticeable. This result was also approved by various studies (ESPAHBODI et al., 2006; BABAAEI et al., 2010;MUJIC et al., 2010) but was in contrast to findings of Bruschi et al. (2003) and Zarafshar et al. (2009). The leaf with code of 1 from group number one showed the maximum Euclidean Distance with the encoded leaf 6 of group number two. These two encoded leaves were both from Shaft region. Thus, it can be realized that biodiversity tends to be high in Shaft which seems to be the result of different micro climate conditions. This result is confirmed by study of Yusefzadeh et al. (2010). The leaves with code of 7 and 9 were both selected from same altitudinal and aspect in Shaft region which showed the minimum Euclidean Distance. In this matter, it can be clarified that the site conditions (e.g. altitudinal and aspect) has important role in formation of groups. Also, this was repeated in sampled fruits of 16 and 17 from Fouman zone with similar site conditions. The scatter plots of leaf and fruit showed that the five specified groups of leaves and the two determined groups of fruits were morphologically similar. Definitely, the leaves and fruits that did not place in any of these groups were quite different. It seems that two factors of altitudinal and aspect are causing the groups segregation. This can be concluded that the differentiations are mainly caused by site conditions changes. This was approved by the study of Alavarez-Alvarez et al. (2006). The grouping is as follows; group 1 with altitude ranges between 400 to 600 meter above sea level, in North to Northwest aspect and average slope of 55 percent, Group 2 with altitude ranges between 280 to 350 meter above sea level, in South to Southwest aspect and average slope of 45 percent and the third group with altitude ranges between 150 to 250 meter above sea level, in Southeast aspect and average slope of 50 percent. Finally, it can be concluded that two important parameters; aspect and altitude highly impacted on the formation of groups, but slope was not remarkable. It is recommended to consider morphological variation in leaf and fruit of sweet chestnut as an important investigation instrument for further genetic diversity studies.
References
- AAS, G., J. MAIER, M. BALTISBERGER , S. METZGER. 1994. Morphology, isozyme variation, cytology, and reproduction of hybrids between Sorbus aria (L.) Crantz and S. torminalis (L.) Crantz. - Botanica Helvetica, 104 (2): 195-214.
- ALDASORO, I.J., E. AEDO, C. NARARRO, F.M. GARMENDIA. 1988. The genus Sorbus ( Malodiaceae, Rosaceae ) in Europe and in North Africa: Morphologyical analyisis systematica. - Systematicatic Botany, 23: 189-212.
- ÁLVAREZ-ÁLVAREZ, P., M. BARRIO-ANTA , U. DIÉGUEZ-ARANDA. 2006. Differentiation of sweet chestnut (Castanea sativa Mill.) cultivars by leaf, nut and burr dimensions. - Forestry, 79 (2): 149-158.
- BABAAEI, F., G. JALALI, D. AZADFAR. 2010. Investigation of genetic variation IZelkova carpinifolia (Pallas) K. Koch, by use of leaf peroxidase isozyme in three lowland habitats in North of Iran. - Journal of Wood and Forest Science and Technology, 19 (1): 121-133.
- BRUSCHI, P., P. GROSSONI, F. BUSSOTTI. 2003. Within- and among-tree variation in leaf morphology of Quercus petraea (Matt.) Liebl. natural populations. - Trees, 17 (2): 164-172.
- ESPAHBODI, K., H. MIRZAEI NADOUSHAN, M. TABARI, M. AKBARINIA , Y. DEHGHANI SHOURAKI. 2006. Investigation of genetic variation of wild service (Sorbus torminalis (L) Crantz), using morphological analysis of fruits and leaves. - Pajouhesh-VA-Sazandegi, 19 (3): 44-57 (In Persian, Summary in English).
- HARRIS, P.J.C., N.M. PASIECZNIK, S.J. SMITH, J.M. BILLINGTON, X. RAMı AMP , L. REZ. 2003. Differentiation of Prosopis juliflora (Sw.) DC. and P. pallida (H. & B. ex. Willd.) H.B.K. using foliar characters and ploidy. - Forest Ecology and Management, 180 (1–3): 153-164.
- HEDAYATI, M. 2003. Study forests of chestnut trees in Gilan and how to multiply and grow Ph.D University of Tehran.
- HEDAYATII, M.A., M.R. MARVI MOHAJER, M.H. DJAZIREIE, M. ZOBEYRI. 2003. An investigation of chestnut Castanea sativa Mill. seedling production in Gilan Province. - Iranian Journal of Natural Resources, 56 (3): 229-244.
- KAFFASH, S., G. BAKHSHI KHAEIKI , B. YOSEFI. 2008. Investigation of leafmorphological characteristics of Quercus infectoria Oliv. (Aleppo Oak) in Kurdestan forests. - Pajouhesh and Sazandegi, 21 (2): 135-144 (In Persian, Summary in English).
- LITTLE, E.L. (1953). Check list of native and naturals, bed trees of the U. S. A. (including Alaska) Washington D. C.: U. S. Forest Service,
- MC DONALD, P.G., C.R. FONSECA., J.M. OVERTON. , M. WESTBOY. 2003. Leaf-size divergence along rainfall and soil-nutrient gradients is the method of size reduction common among clads? - Functional Ecology, 17: 50-57.
- MERWIN, M.L., J.A. MARTIN , R.D. WESTFALL. 1995. Provenance and progeny variation in growth and frost tolerance of Casuarina cunninghamiana in California, USA. - Forest Ecology and Management 79 (3): 161-171.
- MUJIC, I., V. ALIBABICI, J. ZIVKOVIC, S. JAHICI, S. JOKIC, Z. PRGOMET , P. TUZLAK. 2010. Morphological characteristics of chestnut Castanea sativa from the area of NUA- SANA cantion. - Journal of Central European Agriculture 11 (2): 185-190.
- SERDAR, U., N. KURT. 2011. Some Leaf Characteristics are Better Morphometric Discriminators for Chestnut Genotypes. - Journal of Agricultural Science and Technology, 13 (6): 885-894.
- SOKAL, R.R., F.J. ROHLF 1995. Biometry: the principles and practice of statistics in biological research, W.H. Freeman, New York. 413-415 pp
- TAHERI, K. 2011. Wood forest geography, Haghshenas Co. 230 p (In Persian)
- TAHERI, K., B. PILEHVAR 2008. Silviculture, Haghshenas, Tehran. 296 p (In Persian)
- WANG, Y.-F., D.K. FERGUSON, R. ZETTER, T. DENK , G. GARFI. 2001. Leaf architecture and epidermal characters in Zelkova, Ulmaceae. - Botanical Journal of the Linnean Society, 136 (3): 255-265.
- YOSEFZADEH, H., M. TABARI, M. AKBARINIA, M.R. AKBARIAN, F. BUSSOTTI. 2010. Morphological plasticity of Parrotia persica leaves in eastern Hyrcanian forests (Iran) is related to altitude. - Nordic Journal of Botany, 28 (3): 344-349.
- ZARAFSHAR, M., M. AKBARINIA, H. YOSEFZADE., A. SATTARIAN. 2009. Leaf and seed morphological trait in Celtis australis L. in different geographicalcondition. - Tahghighat Genetic, 17: 88-99 (In Persian, Summary in English).



