
International Journal of Plant Science and Ecology, Vol. 1, No. 3, June 2015 Publish Date: Apr. 21, 2015 Pages: 88-92
Screening of Barley Genotypes for Drought Tolerance Using Molecular Markers
M. Huseynova1, *, S. M. Rustamova1, M. Y. Nasrullayeva2, J. A. Aliyev1
1Institute of Botany, Azerbaijan National Academy of Sciences, 40 Badamdar Highway, Baku AZ, Azerbaijan
2Institute of Genetic Reseourses, Azerbaijan National Academy of Sciences, 155 Azadliq Avenue, Baku AZ, Azerbaijan
Abstract
Drought is one of the most important abiotic factors constraining barley production. Drought tolerance of 32 barley (Hordeum vulgare L.) samples of different varieties (21 genotypes-Nutans, 11-Pallidum) and origin have been analyzed with the method of RAPD-PCR. Oligomeric decamer RAPD primers (5′ TCGGCGGTTC 3′) and Р7 (5′ TCGGCGGTTC 3′), associated with drought tolerance have been used. RAPD PCR using P6 marker revealed characteristic loci at 920 bp region in 75% of genotype electrophoretic profiles. Loci were detected at 750 bp region in 78% of the analyzed barley genotypes when using RAPD P7 marker. Comparative analysis of the RAPD spectra showed that characteristic for both markers fragments had been synthesized in 59% of the genotypes (or 19 genotypes). This confirms an existence of a special locus associated with drought tolerance in the barley genotypes. In 6% (2 genotypes) of the studied genotypes characteristic amplification fragments have not been identified with both markers.
Keywords
Hordeum vulgare L., Drought, PCR, RAPD Markers
Received: March 27, 2015
Accepted: April 10, 2015
Published online: April 20, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
1. Introduction
Barley (Hordeum vulgare L.) is one of the earliest global domesticated crop plants in the Neolithic Near East (Salamini et al., 2002). Barley belongs to the grass family Poaceae, the tribe Hordeae and the genus Hordeum. Barley has a large genome of 5.1 gigabases, which was completely sequenced in 2012 (Mayer et al., 2012). Today, in terms of area and production worldwide, barley is the fourth most important cereal after wheat, rice and maize. Approximately 75% of barley global yield is used as feed, 20% as raw material for beverages and 5% as food (Sreenivasulu et al., 2008). Barley is widely adapted to adverse environmental conditions and displays much higher tolerance to different environmental stresses than its close relative wheat (Nevo et al., 2012).
Drought is one of the most important abiotic factors constraining barley production, causing complete grain failure in severely affected fields (Akash, 2013). Improving crop yield under water deficit environments is an urgent task for crop breeding worldwide.
The increasing incidence and importance of drought in relation to crop production has rendered it as a major focus of research for several decades (Budak et al., 2013, Akash, 2013, Ajalli and Salehi, 2012, Altinkut et al., 2003, Bohnert HJ and Jensen., 1996). However, studying drought response is challenged by the complex and quantitative nature of the trait. Drought tolerance is complicated with environmental interactions. In the analysis of a plant’s drought response, the mode, timing, and severity of the dehydration stress and its occurrence with other abiotic and biotic stress factors are significant (Reynolds, 2006). Furthermore, different species, subspecies, and cultivars of crops show variation in their drought tolerance under same conditions, emphasizing the importance of genetic diversity as an underlying factor of drought and its significance in drought-related research. Plants exhibiting high drought tolerance are the most suitable targets of drought-related research and are the most promising sources of drought-related gene and gene regions to be used in the improvement of modern crop varieties (Budak et al., 2013).
In recent years, barley attracts broad attentions as a model crop plant because of its broad natural diversity including geographically diverse elite varieties, landrace and wild accessions and tremendous achievements in barley genomics such as rapid accumulation of EST sequences, extensively characterized genome-wide knockout collections (http://barleygenomics.wsu.edu) and efficient transformation techniques and great efforts on barley genome sequencing and physical mapping (Varshney et al., 2007; Harwood et al., 2009; Mayer et al., 2012). Furthermore, large collections of geographical diverse barley cultivars facilitate to understand the effects of climate changes on crop yield and plant adaptations to environmental stresses.
The utilization of drought tolerant wild species and the rapid advances in molecular biological, functional genomics, and transgenic technologies have facilitated drought-related studies, resulting in significant progress in the identification of related genes and gene regions and dissection of some of its molecular aspects. Molecular breeding approaches based on specific traits utilize molecular markers for the screening of drought tolerance in cultivars. Loci that are targeted in marker-assisted selection (MAS) are most often derived from QTL mapping studies of quantitative traits (Witcombe et al., 2008). Using molecular-genetic methods facilitates to developing new approaches in identification of valuable source material in breeding for heterosis, ensuring reduction of expenses in creation of new competitive hybrids. RAPD marker procedure is simple, fast, does not need previous sequence information, and usually amplify several genetic loci (Akash, 2013).
The aim of the presented work was testing drought tolerance of barley (Hordeum vulgare L.) genotypes of different origin and related to varieties Nutans and Pallidum, using RAPD-markers associated with drought tolerance.
2. Materials and Methods
2.1. Plant Materials
The objects of the study were 32 barley genotypes (Hordeum vulgare L.) of different origin (21 genotypes of Nutans variety and 11 genotypes of Pallidum variety) (Table 2). The plants were cultivated under field conditions at the Galilabad Regional Experimental Station of the Azerbaijan Research Institute of Crop Husbandry. RAPD markers associated with drought tolerance were used for the screening (Pakniyat and Tavakol, 2007).
2.2. DNA Extraction
DNA was extracted from leaves using CTAB method (Murry and Thompson, 1980) with some modifications. Fresh leaf material was cut from the plants and the leaf tissues were ground in a preheated 2×CTAB extraction buffer (100 mM Tris, pH 8, 1.4 M sodium chloride, and 20 mM EDTA, pH 8.0). Liquid nitrogen ground samples were also processed with CTAB buffer. The samples were incubated for 60 minutes in 60° C water bath with occasional vigorous shaking. The samples were mixed gently after adding 400 μl of chloroform and placed on an orbit shaker for 20 minutes at room temperature. After centrifugation at 5 000 rpm, an equal volume of cold absolute isopropanol was added to the supernatant. The solution was well mixed and incubated for 60 minutes at 20° C. The sample was centrifuged for 5 minutes at 5 000 rpm to pellet the DNA followed by washing with 70 % alcohol and then dried at 56 ofor 5 minutes. DNA was resuspended by adding 300 μl TE buffer (10 mM Tris, 1 mM EDTA and pH 8.0).
2.3. DNA Quantification
After diluting the DNA was quantified by taking the optical density (OD) at λ = 260 with a spectrophotometer ULTROSPEC 3300 PRO ("AMERSHAM", USA). The purity of genomic DNA was determined by the A260/A280 absorbance ratio. The quality was also examined by running the extracted DNA samples on 0.8 % agarose gel stained with 10 mg/ml ethidium bromide in 1×TBE (Tris base, Boric acid, EDTA) buffer. The gel was visualized and photographed under UV light.
2.4. Polymerase Chain Reaction Conditions for RAPD
PCR was carried out essentially as described by Williams (Williams et al., 1990). Two 10-mer oligonucleotide primers (Eurogentec S.A., Belgique) were used for DNA amplification (Table 1). Amplifications were performed in «Applied Biosystems 2720 Thermal Cycler» as follows: first 4 min at 94oC followed by 10 cycles of: 1 min at 94oC, 1 min at 36oC and 1 min at 72oC. After that, for next 35 cycles, 0.2oC was added to annealing temperature. After the final cycle, samples were incubated at 72oC for 15 min and then held at 4oC prior to analysis.
Table 1. Primer nucleotide sequence used to amplify DNA

3. Results and Discussion
Drought tolerance of 32 barley (Hordeum vulgare L.) genotypes of different origin has been tested using RAPD-PCR analysis. For this purpose, oligonucleotide decamer RAPD primers-P6 and P7, reported as associated with drought tolerance, were used (Pakniyat and Tavakol, 2007). 21 of these barley genotypes are representatives of Nutans and the rest 11 Pallidum variety.
Figure 1 shows electrophoretic profiles of PCR products obtained with the primer P6 (5′ TCGGCGGTTC 3′). This primer amplifies 920 bp fragments. As seen in table 2 and in the figure expected 920 bp fragments were synthesized in 24 genotypes, which is 75% of the tested genotypes. Despite the fact that they are RAPD primers, they are also associated with a certain locus related to drought tolerance (Pakniyat and Tavakol, 2007).
This indicates that barley genotypes with amplified fragments at 920 bp region have gene loci associated with drought tolerance. As clearly seen in the electrophoretic profiles of 25% of the genotypes (8 genotypes), fragments were not synthesized in the expected region. Five genotypes: St.Garabag 7, Nutans 118/21, (Azerbaijani origin), Zernograd 242, Stepnyak (Russian origin) relate to Nutans variety and the rest three genotypes - K-17893, K-17860, K-7820/2 are of Azerbaijani origin and relate to Pallidum variety.
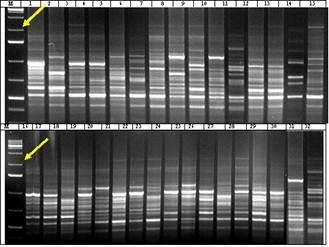
Figure 1. Electrophoretic profiles of Hordeum vulgare L. plants induced with RAPD Р6 (5′ TCGGCGGTTC 3′) primer. Arrows indicate 920 bp region. M (molecular marker)-100 bp. Genotypes: 1 - St.Garabag 7, 2 - Vimpel, 3 - T-78854169, 4 - Strana, 5 - Rabiola, 6 - Zernograd 242, 7 - Polongi, 8 - Ca 56151, 9 - Su 15767, 10 - Rostovskiy738, 11 - Stepnyak, 12 - HW 25325, 13 - Flor 235, 14 - Copelia, 15 - Claret, 16 - Nutans 118/21, 17 - K-90179, 18 - № 77 local, 19 - Nutans 57/9, 20 - Nutans 80-34/14, 21 - Nutans 124/32, 22 - St.Pallidum 596, 23 - T-255/179, 24 - Oglan, 25 - Xemus, 26 - K-2778, 27 - K-7887, 28 - K-818, 29 - K-17893, 30 - K-17860, 31 - K-1783, 32 - K-7820/2.
The second molecular marker used in the study was a primer RAPD P7 (5′ CTGCATCGTG 3′). This primer is responsible for the synthesis of fragments at 750 bp region. Similar to RAPD 6, this primer was also shown to be associated with a locus related to drought tolerance (Pakniyat and Tavakol, 2007).
Figure 2 shows electrophoretic profiles obtained with RAPD P7 primer. Amplification of the fragments at 750 bp region was observed in 25 barley genotypes from 32 ones. Thus, it suggests the presence of a certain gene locus associated with drought tolerance in 78% of the analyzed Hordeum vulgare L. genotypes. As seen in the PCR profiles in 7 genotypes (approximately 22%) 750 bp fragments have not been amplified. They are Zernoqrad 242, Rostovskiy738, Stepnyak genotypes of Russian origin, Flor 235, Copelia genotypes of French origin and K-90179, Nutans 124/32 of Azerbaijani origin. All the genotypes relate to Nutans variety.
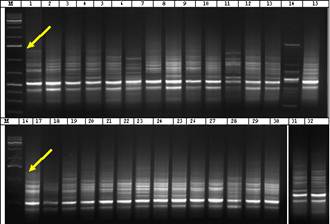
Figure 2. Electrophoretic profiles of Hordeum vulgare L. plants induced with RAPD Р7 (5′ CTGCATCGTG 3′) primer. Arrows indicate 750 bp region. M (molecular marker) – 100 bp. Genotypes: 1 - St.Garabag 7, 2 - Vimpel, 3 - T-78854169, 4 - Strana, 5 - Rabiola, 6 - Zernograd 242, 7 - Polongi, 8 - Ca 56151, 9 - Su 15767, 10 - Rostovskiy738, 11 - Stepnyak, 12 - HW 25325, 13 - Flor 235, 14 - Copelia, 15 - Claret, 16 - Nutans 118/21, 17 - K-90179, 18 - № 77 local, 19 - Nutans 57/9, 20 - Nutans 80-34/14, 21 - Nutans 124/32, 22 - St.Pallidum 596, 23 - T-255/179, 24 - Oglan, 25 - Xemus, 26 - K-2778, 27 - K-7887, 28 - K-818, 29 - K-17893, 30 - K-17860, 31 - K-1783, 32 - K-7820/2.
Comparison of the PCR profiles obtained with RAPD P6 and RAPD P7 primers shows positive results in approximately 59% of the studied barley genotypes in both cases (in 19 from 32 genotypes). So the synthesis of both 920 bp fragments characteristic for P6 primer and 750 bp fragments characteristic for P7 fragments occurred in these genotypes. They are № 77 local, Nutans 57/9, Nutans 80-34/14 genotypes of Azerbaijani origin, Vimpel, Strana,T-78854169 genotypes of Russian origin, HW 25325 of German origin, Polongi of Polish origin, Ca 56151 of Danish origin, Su 15767 of Swiss origin related to Nutans variety and St.Pallidum 596, K-2778, K-7887, K-818, K-1783 of Azerbaijani origin, T-255/179 of Romanian origin, Oglan and Xemus of Bulgarian origin related to Pallidum variety.
Negative results were obtained for only two genotypes, i.e. 6% of the all genotypes for both RAPD primers. Expected fragments with 920 bp and 750 bp sizes are absent among PCR products obtained with both P6 and P7 primers. They are Zernograd 242 and Stepnyak genotypes of Russian origin related to Natans variety.
Table 2. Result of the PCR analysis with Р6 (5′ TCGGCGGTTC 3′) and Р7 (5′ TCGGCGGTTC 3) RAPD markers. [+] – presence of the characteristic locus, ] – absence of the characteristic locus.
| Genotype | Variety | Origin | RAPDP6 | RAPP7 |
| St.Garabag 7 | Nutans | Azerbaijani | - | + |
| Vimpel | Nutans | Russian | + | + |
| T-78854169 | Nutans | Romanian | + | + |
| Strana | Nutans | Russian | + | + |
| Rabiola | Nutans | Polish | - | + |
| Zernograd 242 | Nutans | Russian | - | - |
| Polongi | Nutans | Polish | + | + |
| Ca 56151 | Nutans | Danish | + | + |
| Su 15767 | Nutans | Swiss | + | + |
| Rostovskiy738 | Nutans | Russian | + | - |
| Stepnyak | Nutans | Russian | - | - |
| HW 25325 | Nutans | German | + | + |
| Flor 235 | Nutans | French | + | - |
| Copelia | Nutans | French | + | - |
| Claret | Nutans | English | + | + |
| Nutans 118/21 | Nutans | Azerbaijani | - | + |
| K-90179 | Nutans | Azerbaijani | + | - |
| № 77 local | Nutans | Azerbaijani | + | + |
| Nutans 57/9 | Nutans | Azerbaijani | + | + |
| Nutans 80-34/14 | Nutans | Azerbaijani | + | + |
| Nutans 124/32 | Nutans | Azerbaijani | + | - |
| St.Pallidum596 | Pallidum | Azerbaijani | + | + |
| T-255/179 | Pallidum | Romanian | + | + |
| Oglan | Pallidum | Bulgarian | + | + |
| Xemus | Pallidum | Bulgarian | + | + |
| K-2778 | Pallidum | Azerbaijani | + | + |
| K-7887 | Pallidum | Azerbaijani | + | + |
| K-818 | Pallidum | Azerbaijani | + | + |
| K-17893 | Pallidum | Azerbaijani | - | + |
| K-17860 | Pallidum | Azerbaijani | - | + |
| K-1783 | Pallidum | Azerbaijani | + | + |
| K-7820/2 | Pallidum | Azerbaijani | - | + |
Results obtained with different primers do not match in about 35% of the studied Hordeum vulgare L. genotypes. Thus, for only 5 genotypes from 32 ones (that is about 16%) positive and negative results were obtained for RAPD P6 and RAPD P7 primers, respectively. Electrophoretic profiles obtained with both primers show that 920 bp fragments have been synthesized in five barley genotypes, while the amplification of 750 bp fragments has not occurred. They are Nutans 124/32 and K-90179 genotypes of Azerbaijani origin, Rostovskiy 738 of Russian origin, Flor 235 and Copelia of French origin. It should be noted that all these genotypes relate to Nutans variety. Only for six genotypes (about 19%) of 32 ones positive and negative results were obtained for RAPD P7 and RAPD P6 primers, respectively. Among PCR products of these genotypes, fragments were observed at 750 bp regions contrary to 920 bp. Three of these genotypes St.Garabag 7, Nutans 118/21 and Rabiola are of Azerbaijani origin and relate to Nutans variety. The other three K-17893, K-17860 and K-7820 are of Azerbaijani origin and relate to Pallidum variety.
The study of molecular mechanisms of drought is very important for the creation and selection of plants tolerant to stress (Manavalan, 2009). The complete, systematic and comprehensive study of processes occurring in cereals under drought at molecular –genetic levels is significant for directing stress controlling mechanism and mobilization of all the genetic potential in plants for increasing productivity. The comparative analysis of molecular mechanisms in plants exposed to stress and watered plants allows elucidating such process. Because, genetic potential of cereals are manifested in physiological processes under field conditions. Thus, the choice of experimental materials during the direct selection process and obtaining strong facts for the elucidation of molecular mechanisms of drought tolerance are of great importance for revealing physiological processes occurring at the molecular level in various barley genotypes under water deficiency.
One of the processes occurring in cells under water deficiency is an extensive modification of gene expression leading to the formation of a strong control over the all stress reactions at both physiological and biochemical levels. Specifically, the genes involved in stress reactions have already been identified (Rampino, 2006). Some genes are activated under stress, their protein products are synthesized and joining metabolic pathways they synergistically enhance stress tolerance (Valliyodan & Nguyen, 2006).
It should be noted that in spite of the achievements gained in the field of molecular markers the role of some plant genes in adaptation to environmental stress factors has not been elucidated yet. However, the functions of some genes coding transcription factors and some functional genes coding metabolic components have been studied (Yamaguchi-Shinozaki & Shinozaki 2006; Tran et al., 2007).
Results obtained from RAPD-PCR analysis are a promising start for future studies of plant tolerance to water deficiency. Investigations carried out at molecular-genetic level allow revealing specific genetic markers for the selection of genotypes tolerant to adverse environmental factors. Purposeful use of specific primers allows testing drought tolerance of a lot of wheat genotypes in a short period of time saving intellectual power of researchers and experimental tools. Owing to RAPD-PCR technology it is possible to differentiate markers related to drought tolerance in cereals and all the effects leading to genetic variations of barley genotypes. Markers based on genotypes will contribute to future increase of the effectiveness in barley selection.
References
- Ajalli J. and Salehi M. (2012) Evaluation of drought stress indices in barley (Hordeum vulgare L.). Ann. Biol. Res., 3: 5515-5520.
- Akash M.W. (2013) Development of SCAR markers for molecular tagging of drought tolerance QTL in barley. Life Sci. J., 10(12s).
- Altinkut A., Kazan K., Gozukirmizi N. (2003) AFLP marker linked to water-stress-tolerant bulks in barely (Hordeum vulgare L.). Genet. Mol. Biol., 26(1): 77-82.
- Bohnert H.J. and Jensen R.G. (1996) Strategies for engineering waterstress tolerance in plants. Trends in Biotech., 14: 89-97.
- Budak H., Akpinar B.A., Unver T., Turktas M. (2013) Proteome changes in wild and modern wheat leaves upon drought stress by two-dimensional electrophoresis and nanolc-esi–ms/ms. Plant Mol. Biol. : 1-15.
- Harwood V.J., Brownell M., Wang S., Lepo J., Ellender R.D., Ajidahun A., Hellein K.N., Kennedy E., Ye X., Flood C. (2009). Validation and field testing of libraryindependent microbial source tracking methods in the Gulf of Mexico. Water Res., 43(19): 4812–9.
- Manavalan L.P., Guttikonda S.K., Tran L.S., Nguyen H.T. (2009) Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol., 50(7):1260-1276.
- Mayer K.F., Waugh R., Brown J.W., Schulman A., Langridge P., Platzer M., Fincher G.B., Muehlbauer G.J., Sato K., Close T.J., Wise R.P., Stein N. (2012) A physical, genetic and functional sequence assembly of the barley genome. Nature, 491:711-716.
- Murray M.G., Thompson W.F. (1980) Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res., 8:4321-4325.
- Nevo E., Fu Y.B., Pavlicek T., Khalifa S., Tavasi M., Beiles A. (2012) Evolution of wild cereals during 28 years of global warming in Israel. Proc. Natl. Acad. Sci. USA, 109:3412-5. doi: 10.1073/pnas.1121411109.
- Pakniyat H., Tavakol E. (2007) RAPD markers associated with drought tolerance in bread wheat (Triticum aestivum L.). Pakistan J. Biol. Sci., 10: 3237-3239.
- Rampino P., Pataleo S., Gerardi C., Perotta C. (2006) Drought stress responses in wheat: physiological and molecular analysis of resistant and sensitive genotypes. Plant Cell Environ, 29: 2143-2152.
- Reynolds M. P. (2006) "Drought adaptation in wheat," in Drought Tolerance in Cereals. J. M. Ribaut, Ed., chapter 11, pp. 402-436, Haworth’s Food Products Press, New York, NY, USA.
- Salamini F., Ozkan H., Brandolini A., Schafer-Pregl R., Martin W. (2002) Genetics and geography of wild cereal domestication in the Near East. Nat. Rev. Genet., 3: 429–441.
- Sreenivasulu N., Usadel B., Winter A., Radchuk V., Scholz ., Stein N., Barley grain maturation and germination: metabolic pathway and regulatory network commonalities and differences highlighted by new MapMan/PageMan profiling tools. Plant physiology, 146(4):1738-1758.
- Tran L.S., Nakashima K., Shinozaki K., Yamaguchi-Shinozaki K. (2007) Plant gene networks in osmotic stress response: from genes to regulatory networks. Meth. in Enzymol., 428: 109-128.
- Valliyodan B. and Nguyen H.T.(2006)Understanding regulatorynetworks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol., 9: 189-195.
- Varshney R.K., Beier U., Khlestkina E., Kota R., Korzun V., Röder M., Graner A., Börner A. (2007) Single nucleotide polymorphisms in rye: discovery, frequency and applications for genome mapping and diversity studies. Theoret. and Appl. Gen., 114: 1105-1116.
- Williams J.G., Kubelik K.J., Livak J.A., Tingey S.V. (1990) DNA polymorphisms amplified by arbitrary primers are useful genetic markers. Nucleic Acids Res., 18: 6531-6535.
- WitcombeJ.R, HollingtonP.A, HowarthC.J,ReaderS,SteeleK.A.(2008)Breeding for abiotic stresses for sustainable agriculture.Phil. Trans. R. Soc. B.,363:703-716.
- Yamaguchi-Shinozaki K., Shinozaki K. (2006) Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol., 57: 781-803.



