
International Journal of Animal Biology, Vol. 1, No. 5, October 2015 Publish Date: Jul. 28, 2015 Pages: 231-236
Effects of D-Galactose Induced Oxidative Stress and Ethanolic Extracts of Bacopa monniera and Lactuca sativa on Non-Specific Esterase in Midgut of Silkworm Bombyx mori
Y. B. Gaikwad1, J. A. Chavan1, G. P. Bhawane1, *, N. K. Desai2
1Department of Zoology, Shivaji University, Kolhapur, M. S. India
2Department of Chemistry, Shivaji University, Kolhapur, M. S. India
Abstract
The non-specific esterase (NSE) activity was estimated in the midgut of simultaneously and independently treated larvae of silkworm Bombyx mori (Race-PM) with D-galactose and ethanolic extract of Bacopa monniera and Lactuca sativa. Treatments of only D-galactose showed non-significant (p > 0.05) decrease in NSE activity as compared with control group. Concomitant treatment of D-galactose and ethanolic extract of B. monniera and L. sativa showed non-significant change in non-specific esterase activity by as compared with control group. There was significant (p < 0.05) change in NSE activity in the groups treated with only ethanolic extract of B. monniera and L. sativa. The lipid peroxidation and fluorescence was measured to analyze the extent of oxidative stress. The results showed high degree of positive correlation (r =0.8088) between oxidative stress and non-specific esterase activity.
Keywords
Lipid Peroxidation, Fluorescence, Aging and Correlation
Received: June 19, 2015
Accepted: July 10, 2015
Published online: July 27, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
Contents
1. Introduction 2. Material and Methods 2.1. Rearing of Silkworm 2.2. Preparation of Ethanolic Extract 2.3. Administration of D-Galactose and Ethanolic Extracts 2.4. Estimation of Lipid Peroxidation 2.5. Spectrofluorometric Analysis 2.6. Estimation of NSE 2.7. Estimation of Protein 2.8. Statistical Analysis 3. Results and Discussion Acknowledgement
1. Introduction
Non-specific esterase (NSE) is a lysosomal enzyme and is co-localized in primary lysosomes and in a large diverse population of autophagic vacuoles (Willard et al., 1984; Jimenez and Gilliam, 1990). NSE enzymes are believed to participate in the intracellular hydrolysis of the ingested material (Bang, 1975; Cheng et al., 1974; Michelson, 1975). The nonspecific esterases are broadly distributed enzymes that catalyze the hydrolysis of ester and amide linkages. They fall into several different classes which can be distinguished by their substrate preferences and their sensitivity to inhibitors.
Esterase enzymes play an important role in conferring or contributing to insecticide resistance in insects. This has been shown in aphid Myzus persiace (Field and Devonshire, 1998), mosquitoes, Culex quinquefasciatus and C. pipiens (Guillemaud et al., 1997), blowfly, Lucilia cuprina (Campbell et al., 1998) and housefly Musca domestica (Claudianos et al., 1999; Taskin and Kence, 2004). The biochemical and physiological properties of esterases were previously studied (Healy et al., 1991; Oakeshott et al., 1993). In insects, the esterase enzyme patterns have shown high rates of intraspecific and interspecific variations (Nascimento and Bicudo, 2002).
Possibly, gene duplication followed by divergence of duplicated genes from the ancestral gene is the origin of at least part of this variability (Oakeshott et al., 1993; Nascimento and Bicudo, 2002). Acetylcholinesterase enzyme, a key enzyme in the insect central nervous system, terminates nerve impulses by catalyzing the hydrolysis of the neurotransmitter acetylcholine. The resistance associated with modification of acetylcholinesterase makes it less sensitive to inhibition by organophosphates and carbamate insecticides (Feyereisen, 1995). Several point mutations have been identified in D. melanogaster (Mutero et al., 1994; Menozzi et al., 2004) and M. domestica Ace (acetyl choline esterase) genes, encoding acetyl choline esterase (Kozaki et al., 2001).
In insects, esterases are involved in important physiological processes, including the catabolism of juvenile hormone (Zera et al., 1992, 2002), pesticide resistance (Whyard et al., 1995; Rosario-Cruz et al., 1997), digestion (Kerlin and Hughes, 1992; Argentine and James, 1995) and reproduction (Richmond and Senior, 1991; Karotam and Oakeshott, 1993). Esterases are main hydrolytic enzymes responsible for converting complex storage lipids into easily utilizable glycerides (Subramaniam, 1982).
Oxidative stress is caused by the imbalance between the reactive oxygen species production and elimination in the biological system. D-galactose is a naturally occurring sugar in the body; however, at high levels, it can cause the accumulation of reactive oxygen species, finally resulting in increased oxidative stress (Shan et al., 2009). D-galactose has been used to inject mice or rats for pharmacological studies. There are several evidences which shows that D-galactose cause aging-related changes including the increase of reactive oxygen species and the decrease of antioxidant enzyme activity (Liu et al., 2009; Wang et al., 2009). Administration of D-galactose showed increased lipid peroxidation and fluorescence, decreased antioxidant enzyme activity and reduced life span (Cui et al., 2004).
The alcoholic extract of Bacopa monniera (Bramhi) has been shown to be a potent antioxidant, free radical scavenger and anti-lipid peroxidative agent (Gajare et al., 2006; Bhattacharya et al., 2000). B. monniera contains tetracyclic triterpenoid saponins, bacosides A and B, hersaponin, alkaloids like herpestine, brahmine and flavonoids (Kirtikar and Basu, 1994). Ethanolic extract of Lactuca sativa (Lettuce) contains quercetin which is a potent antioxidant flavonoid. Quercetin showed increased antioxidant enzyme activity of mice brain (Su et al., 2007). Treatments of ethanolic extracts of B. monniera and L. sativa showed reduced lipid peroxidation and increased membrane integrity in silkworm B. mori (Gaikwad et al., 2010).
There is scanty literature available regarding the role of NSE during aging. Therefore, in present work simultaneous and independent treatment of D-galactose, ethanolic extracts of B. monniera and L. sativa in midgut of silkworm B. mori to study the pattern of NSE activity during aging process.
2. Material and Methods
2.1. Rearing of Silkworm
The silkworm larvae of race PM was reared in laboratory conditions at 280C and 80% humidity as suggested by Krishnaswami et al., (1978).
2.2. Preparation of Ethanolic Extract
The leaves of B. monniera and L. sativa were washed with distilled water and shade dried for 3 to 4 days. After complete drying the leaves were powdered and kept in ethanol for 72 hours. The alcohol was allowed to evaporate and extract was collected and stored in refrigerator for further use.
2.3. Administration of D-Galactose and Ethanolic Extracts
The larvae were divided into 6 groups containing 30 larvae in each group. The treatment was given during first three days of forth and fifth instars. Mulberry leaves of equal weight were dipped in various concentrations of D-galactose and ethanolic plant extracts (Table - 1) and air-dried. The larvae were fed on the dried leaves once in a day. On the forth day of fifth instar the midgut was removed and homogenized in 0.8% NaCl. Homogenate was centrifuged at 1000 g and supernatant was used as sample.
2.4. Estimation of Lipid Peroxidation
The lipid peroxidation was estimated by method of Buege and Aust (1987). The reaction mixture contained 1 ml of sample and 2 ml TCA-TBA-HCl reagent (15 % TCA + 0.375% TBA in 0.25M HCl). The tubes were kept in boiling water bath for 10 min. cooled and centrifuged. The absorbance of supernatant was measured at 532 nm on spectrophotometer. The amount of malondialdehyde (MDA) was calculated using extinction coefficient 1.56X105/M MDA/cm2.
2.5. Spectrofluorometric Analysis
Spectroflurometric analysis was performed by method of Dillard and Tappel (1984). 1 ml of tissue sample was taken in clean dry test tube. 6 ml of chloroform methanol mixture was added and the tubes were shaken for proper mixing. The mixture was centrifuged for 10 in at 1000g. 6 ml distilled water was added slowly in test tube and 4 ml of upper chloroform layer was removed to a test tube and 0.4 ml methanol was added. The fluorescence was determined on spectroflurometer and the results were expressed in intensity/mg protein.
2.6. Estimation of NSE
The NSE activity from sample was estimated by method described by Subramaniam (1982). The tissues were homogenized in 0.8% saline and centrifuged at 1000g for 10 min. The pellet was discarded and supernatant (sample) was used for estimation of NSE activity. Assay mixture consisted of 0.5 ml tissue sample, 2.5 ml of substrate buffer containing 600 µg/ml α-naphthyl acetate in 50 mM phosphate buffer (pH 7.0), tubes were incubated at 40oC for 20 min. Finally 2.0 ml of arrest regent (1% fast blue RR and 1% sodium dodecyl sulfate in 1:5 proportion) was added. Blank tube contained 0.5 ml of distilled water instead of tissue homogenate and arrest reagent was added immediately. Assay control tube contained similar composition as blank but was incubated along with sample tubes to avoid increase in absorbance due to auto-hydrolysis of α-naphthyl acetate. Finally absorbance of control was subtracted from sample absorbance. The NSE activity was expressed as µg α-naphthol/mg protein/hr.
2.7. Estimation of Protein
The soluble protein from sample was estimated by Lowry’s method (Lowry et al., 1951).
2.8. Statistical Analysis
Kruskal-Wallis test was implemented for data analysis and Karl Pearson’s correlation coefficient method was used to study correlation between lipid peroxidation and NSE activity.
Table 1. Groups and treatments given.
| Group | Treatments | Group | Treatments |
| Control | Distilled water | 30L | 30 mg/ml D-galactose + 2 mg/ml L. sativa |
| 30D | 30 mg/ml D-galactose | B | 2 mg/ml B. monniera |
| 30B | 30 mg/ml D-galactose + 2 mg/ml B. monniera | L | 2 mg/ml L. sativa |
3. Results and Discussion
D-galactose is a reducing sugar and can be metabolized at normal concentration. However, at high levels, it is converted into aldose and hydroperoxide under the catalysis of galactose oxidase, resulting in the generation of a superoxide anion and oxygen-derived free radicals (Wu et al., 2008). In insects enzymes of defense mechanism includes superoxide dismutase, catalase, ascorbate peroxidase while, non-enzyme molecules are α-tocopherol, ascorbic acid and glutathione besides these dietary flavonoids also play crucial role in nullifying reactive oxygen species. Overwhelming reactive oxygen species cause deleterious effects on biological molecules resulting in lipid peroxidation, protein oxidation, DNA damage and lipofuscinogenesis (Sohal et al., 1995; Standtman, 1992, Finkel and Hallbrook, 2000).
Lipid peroxidation is one of the major manifestations of aging. In present investigation the lipid peroxidation was measured in midgut tissue for determination of extent of oxidative stress caused by D-galactose treatment. Results showed highly significant (p < 0.001) increase in the lipid peroxidation of D-galactose treated group as compared with the control group (Fig. 1). The simultaneous treatment of D-galactose and ethanolic extract of B. monniera showed decreased lipid peroxidation as compared with only D-galactose treated group showing protective effect of ethanolic extract of B. monniera. There was non-significant (p > 0.05) decrease in the group treated with D-galactose and ethanolic exctract of L. sativa as compared with the control group. The groups treated with only ethanolic extracts of B. monniera and L. sativa showed highly significant and moderately significant (p < 0.01) decrease in lipid peroxidation respectively as compared with control group.
Ethanolic extract of B. monniera contains number of antioxidant flavonoids like triterpenoid glycosides, bacosides and flavonol which are proven to have antioxidant properties (Chillara et al., 2005). Previous studies have detected significant levels of antioxidant activities and phenolic components in L. sativa (Caldwell, 2003; Cao et al., 1996; Chu et al., 2002; Vinson et al., 1998). The antioxidant extracts of L. sativa exhibited significant oxygen radical absorbance capacity (Caldwell, 2003; Cao et al., 1996), prevention of lipid oxidation in lower-density lipoproteins (Vinson et al., 1998), inhibitory effects on ethylene formation induced by oxyradicals (Chu et al., 2002), and hydroxyl radical absorbance capacity and inhibitory activity against Cu++induced protein oxidation.
Lipofuscinogenesis is unavoidable process which progresses with aging. Lipofuscin granules accumulate in postmitotic cells causing hurdles in cellular processes. The lipofuscin granules contain non-degradable fluorescent products. The spectrofluorometric analysis of midgut homogenates showed excitation wavelength 360 nm and emission wavelength 524 nm. The intensity of fluorescence was higher in the midgut tissue of D-galactose treated group as compared with the control group (Fig. 2). The intensity of fluorescent products was low in the group treated with ethanolic extract of B. monniera and ethanolic extract of L. sativa treated groups showing their antioxidant properties. In earlier studies Kalamade et al., (2008) reported similar decrease in lipofuscinogenesis in mice brain when treated with ethanolic extract of B. monniera. Similarly ethanolic extract of L. sativa also showed decreased lipofuscin granules in nervous tissue as well as midgut. L. sativa extract contain quercetin which is reported by several authors as potent antioxidant (Havsteen, 2002).
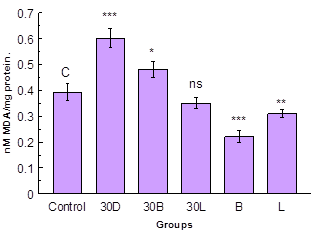
Fig. 1. Lipid peroxidation in midgut.
Data are Mean ± S.D. (n = 3); *, **, *** and ns indicates significance
level p < 0.05, p < 0.01, p < 0.001 and p > 0.05 respectively.

Fig. 2. Fluorescence in midgut.
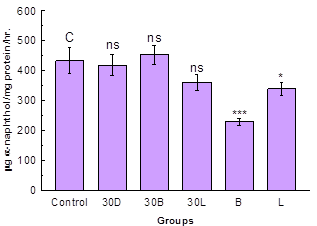
Fig. 3. NSE activity in midgut.
Data are Mean ± S.D. (n = 3); *, *** and ns indicates significance level p < 0.05, p < 0.001 and p > 0.05 respectively
The NSE activity was decreased in the midgut of D-galactose treated group and the group treated simultaneously with D-galactose and ethanolic extract of L. sativa however the decrease was non-significant (p > 0.05) as compared with control group (Fig. 3). In the group treated simultaneously with D-galactose and ethanolic extract of B. monniera showed non-significant increase in NSE activity as compared with control group. Treatment with only ethanolic extracts of B. monniera and L. sativa showed highly significant (p < 0.001) and moderately significant (p < 0.01) decrease in NSE activity as compared with control groups respectively. Statistical analysis for correlation between lipid peroxidation and NSE activity showed high degree of positive correlation (r =0.8088).
Juvenile hormones are a group of acyclic sequiterpenoids that regulate many aspects of insect physiology. Juvenile hormones play crucial role in development, reproduction diapause and polyphenisms. NSE plays an important role in the metamorphosis of an insect by complete degradation of juvenile hormones (Riddiford, 1994; Wyatt and Davey, 1996; Nijhout, 1994). In Drosophila melanogaster 8 to 16% decrease in esterase activity was observed when the activity was measured on 10, 25 and 57 day of life span (Burcombe, 1972), while in Callosobruchus maculatus NSE activity was found to increase throughout life (Sharma and Sharma, 1981). Our results show there is increase in NSE activity as aging advances.
Acknowledgement
Author Y. B. Gaikwad is thankful to University Grants Commission, New Delhi for providing Meritorious Research Fellowship.
References
- Argentine, J.A. and James, A.A. (1995). Characterization of a salivary gland-specific esterase in the vector mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 25, 621–630.
- Bang F. B. (1975). Phagocytosis in invertebrates. In Maramorosch, K. and Shope, R. E., eds., Invertebrate Immunity, Academic Press, New York, pp. 137-151.
- Bhattacharya, S.K., Bhattacharya, A., Kumar, A., Ghosal, S. (2000). Antioxidant activity of B. monniera in rat frontal cortex, striatum and hippocampus. Phytotherapy Research 14, 174–179.
- Buege, J. A. and Aust, S. D. (1987). Microsomal lipid peroxidation. Methods Enzymol. 52, 302–310.
- Caldwell, C. R. (2003). Alkylperoxyl radical scavenging activity of red leaf lettuce (Lactuca sativa L.) phenolics. Journal of Agricultural and Food Chemistry, 51, 4589–4595.
- Campbell P. M., Yen J. L. and Masoumi A. (1998). Cross-resistance patterns among Lucilia cuprina (Diptera: Calliphoridae) resistant to organophosphorus insecticides. J Economic Entomol. 91(2): 367-375, 1998.
- Cao, G., Sofic, E. and Prior, R. L. (1996). Antioxidant capacity of tea and common vegetables. Journal of Agricultural and Food Chemistry , 44, 3426–3431.
- Cheng T. C., Cali A. and Foley D. A. (1974). Cellular reactions in marine pelecypods as a factor influencing endosymbioses. In Vernberg, W. B., ed., Symhosis in the Sea, Univ. South Carolina Press, Columbia, pp. 62-91.
- Chillara Sivaramakrishna, Rao Chirravuri V., Golakoti Trimurtulu, Mulabagal Vanisreeb, Gottumukkala V. Subbaraju (2005).Triterpenoid glycosides from Bacopa monnieri. Phytochemistry 66 (2005) 2719–2728.
- Chu, Y. F., Sun, J., Wu, X. and Liu, R. H. (2002). Antioxidant and antiproliferative activities of common vegetables. Journal of Agricultural and Food Chemistry, 50, 6910–6916.
- Claudianos C, Russell R. J. and Oakeshott J. G. (1999). The same amino acid substitution in orthologous esterases confers organophosphate resistance on the house fly and a blowfly. Insect Biochemistry and Molecular Biology. 29(8): 675-686, 1999.
- Cui X, Wang L, Zuo P, Han Z, Fang Z, Li W, Liu J. (2004).D-galactose-caused life shortening in Drosophila melanogaster and Musca domestica is associated with oxidative stress. Biogerontology. 2004; 5(5):317-25.
- Dillard Cora J.and Tappel A. L.(1984). Fluorescent damage products of lipid peroxidation. Methods in Enzymology, Volume 105, 1984, Pages 337–341.
- Feyereisen R. (1995). Molecular biology of insecticide resistance. T oxicology Letters. 82(3): 83-90, 1995.
- Field, L. M. and Devonshire, A. L. (1998). Evidence that the E4 and FE4 esterase genes responsible for insecticide resistance in the aphid Myzus persicae (Sulzer) are part of a gene family. Biochem. J. 330, 169-173.
- Finkel Toren and Holbrook Nikki J. (2000). Oxidants, oxidative stress and the biology of ageing. Nature, Vol 408, 239 – 247.
- Gaikwad Y. B., Gaikwad S. M., Bhawane GP (2010) Effect of induced oxidative stress and herbal extracts on acid phosphatase activity in lysosomal and microsomal fractions of midgut tissue of silkworm Bombyx mori. Journal of insect science, Volume - 10, 1 -9.
- Gajare, K.A., Deshmukh, A.A. and Pillai M.M. (2007). Neuroprotective effect of Bacopa monniera Linn. Extract on lipofuscinogenesis and fluorescence product in the brain of D-galactose induced ageing accelerated mice. J. Cell Tissue Res.7 (2): 1167-1172, (2007).
- Guillemaud, T., Makate, N., Raymond, M., Hirst, B. and Callaghan, A. (1997) Esterase gene ampli¢cation in Culex pipiens. Insect Molec. Biol. 6, 319-327.
- Havsteen B.H. (2002). The biochemistry and medical significance of flavonoids. Pharmacology and therapeutics 96 (2002), 67-202.
- Healy, M. J., Dumancic, M. M., and Oakeshott, J. G. (1991). Biochemical and physiological studies of soluble esterases from Drosophila melanogaster.Biochem. Genet. 29:365.
- Jimenez D.R. and Gilliam M. (1990). Ultrastructure of the ventriculus of the honey bee, Apis mellifera (LJ: Cytochemical localization of acid phosphatase, alkaline phosphatase, and nonspecific esterase. Cell Tissue Res 261:431-443.
- Kalamade V. I., Pillai M.M. and Kalamade I. S. (2008).Effect of Bacopa monniera (Linn.) on lipid peroxidation and lipofuscinogenesis in prostate gland of D-galactose induced aging mice, Mus musculus. Indian J Exp Biol. 2008;46 (7): 547-549.
- Karotam, J. and Oakeshott, J. G. (1993). Regulatory aspects of esterase 6 activity variation in sibling Drosophila species. Heredity 71, 41–50.
- Kerlin, R.L. and Hughes, S. (1992). Enzymes in saliva from four parasitic arthropods.Med. Vet. Entomol. 6, 121–126.
- Kirtikar K. R. and Basu B. D. (1994). Indian Medicinal plants. Vol I, 1816-1817.
- Kozaki T., Shono T., Tomita T. and Kono Y. (2001).Fenitroxon insensitive acetylcholinesterases of the housefly, Musca domestica, associated with point mutations. Insect Biochem Mol Biol. 31: 991-997, 2001.
- Liu A., Ma Y., and Zhu Z. (2009).Protective effect of selenoarginine against oxidative stress in D-galactose-induced aging mice, Bioscience, Biotechnology and Biochemistry, vol. 73, no. 7, pp. 1461–1464.
- Lowry, O. R., Rosebrow, N. J. and Randall, R. J. (1951). Protein measurement with the folin phenol reagent. Biol. Chem. 193, 265 – 275.
- Krishnaswami S., Narsimhana N., Suryanarayana S.K., Kumararaji S. (1978). Silkworm rearing, Manual on Sericulture Vol.2. FAO. Agricultural servicesBulletin. No. 15.
- Menozzi P., Shi M. A., Lougarre A., Tang Z. H. and Fournier D. (2004). Mutations of acetylcholinesterase which confer insecticide resistance in Drosophila melanogaster populations. BMC Evolutionary Biology 4: 4.
- Michelson E. H. (1975). Cellular defense mechanisms and tissue alterations in gastropod molluscs. In Maramorosch, K. and Shope, R. E., eds., lnvertebrate Immunity, Academic Press, New York, pp. 181-195.
- Mutero A., Pralavorio M., Bride J. M. and Fournier D. (1994) Resistance-associated point mutations in insecticide-insensitive acetylcholinesterase. Proceedings of the national academy of sciences 91: 5922–5926.
- Nascimento A.P. and Bicudo H.E.M.D. (2002). Esterase patterns and phylogenetic relationships of Drosophila species in the saltans subgroup (saltans group). Genetica. 114(1): 41-51, 2002.
- Nijhout, H. F. (1994). Insect Hormones. Princeton University Press, Princeton.
- Oakeshott, J. G., van Papenrecht E.A., Boyce T. M., Russell R.J. and Healy M. J. (1993). Evolutionary genetics of Drosophila esterases. Genetica. 90: 239-268, 1993.
- Qun Shan, Jun Lu, Yuanlin Zheng, Jing Li, Zhong Zhou, Bin Hu, Zifeng Zhang, Shaohua Fan, Zhen Mao, Yong-jian Wang and Daifu Ma. (2009). Purple sweet potato color ameliorates cognition deficits and attenuates oxidative damage and inflammation in aging mouse brain induced by D-Galactose. Hindawi Publishing Corporation Journal of Biomedicine and Biotechnology, Volume 2009, Article ID 564737, 1 -9.
- Richmond, R. C. and Senior, A. (1991). Esterase 6 of Drosophila melanogaster: Kinetics transfer to females, decay in females and males recovery. J. Insect Physiol. 27, 849–853.
- Riddiford, L. M. (1994). Cellular and molecular actions of juvenile hormone. I. General considerations and premetamorphic actions. Adv. Insect Physiol. 24, 213-274.
- Rosario-Cruz, R., Miranda-Miranda E., Garcia-Vasquez Z. and Ortiz-Estrada M. (1997).Detection of esterase activity in susceptible and organophosphate resistant strains of the cattle tick Boophilus microplus (Acari: Ixodidae). Bull. Entomol. Res. 87, 197–202.
- Shan Q. Lu J. Zheng Y. Li J. Zhou Z. Hu B. Zhang Z. Fan S. Mao Z. Wang YJ. Ma D. Purple sweet potato color ameliorates cognition deficits and attenuates oxidative damage and inflammation in aging mouse brain induced by d-galactose. J Biomed Biotechnol. 2009 doi: 10.1155/2009/564737.
- Sharma, G. and Sharma, S. P. (1981). Age dependent changes in esterases of Callosbruchus maculatus Fab. (Bruchidae: Coleoplerii). Exp. Aging Res, 7. 107-115.
- Sohal, R. S., S. Agarwal, and B. H. Sohal (1995). Oxidative stress and aging in the Mongolian gerbil (Meriones unguiculatus).Mech. Ageing Dev. 81: 15 – 25, 1995.
- Standtman E. R. (1992). Protein oxidation and aging. Science, 257: 1220 –1224.
- Su Wen Sun, Hu Qing Yu, Hong Zhang, Yuan Lin Zheng, Jian Jun Wang and Lan Luo (2007). Quercetin attenuates spontaneous behavior and spatial memory impairment ind-galactose–treated mice by increasing brain antioxidant capacity. Nutrition Research 27 (2007) 169–175.
- Subramamiam T. (1982) Quantitative assay of non-specific esterase in the developing egg of Emerita asiatica. Manual for research methods for marine invertebrate reproduction. CMFRI special publication No. 9.
- Taskin V. and Kence M. (2004). The genetic basis of malathion resistance in housefly (Musca domestica L.) strains from Turkey. Russian Journal of Genetics. 40: 1475-1482, 2004.
- Vinson, J. A., Hao, Y., Su, X. and Zubik, L. (1998). Phenol antioxidant quantity and quality in foods: Vegetables. Journal of Agricultural and Food Chemistry, 46, 3630–3634.
- Wang W., Li S., Dong H.-P., Lv S., and Tang Y.Y. (2009). Differential impairment of spatial and nonspatial cognition in a mouse model of brain aging, Life Sciences, vol. 85, no. 3-4, pp. 127– 135.
- Whyard, S., Down, A. E. R. and Walker, V. K. (1995). Characteriza-tion of a novel esterase conferring insecticide resistance in the mosquito Culex tarsalis. Arch. Insect Biochem. Physiol. 29, 329–342.
- Willard O. Granath, Jr.; Timothy and P. Yoshino. (1984). Intracellular Distribution of Lysosomal Enzymes within the Hemocytes of Biomphalaria glabrata. Transactions of the American Microscopical Society, Vol. 103, No. 1. (Jan., 1984), pp. 38-43.
- Wu D., Lu J., Zheng Y., Z. Zhou, Q. Shan, and D. F. Ma (2008.). Purple sweet potato color repairs D-galactose-induced spatial learning and memory impairment by regulating the expression of synaptic proteins. Neurobiology of Learning and Memory, vol. 90, no. 1, pp. 19–27.
- Wu, D.J. and Hou, R.F. (1993). Relationship between thermotolerancy and heat-stable esterase in the silkworm Bombyx mori L. (Lepidoptera: Bombycidae). Appl. Entomol. Zool. 28, 371–377.
- Wyatt G.R. and Davey K.G. (1996). Cellular and molecular actions of juvenile hor-mone. II. Roles of juvenile hormone in adult insects. Adv. Insect Physiol. 26, 1-155.
- Zera, A. J., T. Sanger, J. Hanes, and L. Harshman. (2002). Purification and characterization of hemolymph juvenile hormone esterase from the cricket, Gryllus assimilis. Arch. Insect Biochem. Physiol, 4941-55.
- Zera, A.J., Gu, X. and Zeisset M. (1992). Characterization of juvenile hormone esterase from genetically-determined wing morphs of the cricket Gryllus rubens. Insect Biochem. Mol. Biol. 22, 829–839.



