
International Journal of Animal Biology, Vol. 1, No. 2, April 2015 Publish Date: Mar. 21, 2015 Pages: 17-27
The Analysis of Phrase Combinations and Various Ways of Syllabic Singing at Vocal Variability of Song Types of Chaffinch (Fringilla coelebs L.)
O. A. Astakhova*
Moscow State University of M. V. Lomonosov, Biology Faculty, Department of Vertebrate Zoology, Moscow, Russia
Abstract
Questions of evolutionary development of vocalizations (songs, calls) in time and space of many species of birds already for a long time are mentioned in bioacoustics. Connection of geographical variability and structural features of vocalizations was one of the first preconditions of evolutionary hypotheses. Development of song cultures at species of birds with the big repertoires, having social training traditions, remains till now in many respects not clear. In article some features of formation of new song types of chaffinch (Fringilla coelebs L.) and their territorial distribution are considered.
Keywords
Birds Song, Species-specific Song, Parts (Phrases) of Song, Phonetic Distinctions, Lexical Distinctions
Received: March 5, 2015
Accepted: March 16, 2015
Published online: March 20, 2015
@ 2015 The Authors. Published by American Institute of Science. This Open Access article is under the CC BY-NC license. http://creativecommons.org/licenses/by-nc/4.0/
Contents
1. Introduction 2. Material and Methods 3. Results and Discussion 3.1. Replacement of a Phrase of One Kind by Another in Songs of One Type at Singing 3.2. Addition of a Phrase to Songs of One Type at Singing 3.3. Removal of a Phrase from Songs of One Type at Singing 3.4. Phrase Combinations of Different Song Types at Singing 3.5. Syllabic Variability in Started Singing of Song Types 3.6. Syllabic Variability of Elements in Trill of Song Types 3.7. Syllabic Variability in Final Stroke of Chaffinch Song Types (Fringilla coelebs L.) 4. Conclusion
1. Introduction
Explanation of birds song variability shows, that traditional patterns of vocal behaviour do not exist equally with morphological features (more likely, as separate aspect). It partly due result of cultural evolution, which represents the important factor – as though a traditional heredity is developed, but, apparently, it not a parameter in morphological variability (Lemon, 1975; Slater, Ince, 1979; Mundinger, 1980).
Basic (base) structure of birds vocalization represents fundamental, stabilized, species specific characteristic feature of vocalization (Mundinger, 1979). Many local populations of birds are characterized by the population specific image (on sonogram) of a vocal pattern (dialect), which has certain lexical (dictionary structure), morphological (an accent, structure) and phonetic (pronunciation) features (Kurath, 1972).
Thielcke G. (1965, 1969) defined dialects of birds as vocal variants with mosaic distribution. This mosaic definition one of the basic, which are widely used, but the majority of researchers of microgeographical variability not precisely enough assert in studying vocal distribution, whether there is a mosaic pattern (Kreutzer, 1974; Kroodsma, 1974).
Regional vocal variability of birds can be determined in qualitative aspect (the form of a syllable, syntax) and in quantitative parameters (frequency - KHz, time parameters - sec, min). For many species specific patterns of variability as purchases (having) of vocal traditions are products of cultural evolution. Cultural evolution is a parameter for species of birds with a historical variety of patterns of microgeographical variability, and also for species with differentiated vocal syllables (Mundinger, 1980).
Many characteristics of a vocal of birds are precisely mentioned (related) by social training traditions (song learning), receiving their vocal patterns by specific imitation. Therefore the tradition, custom should be taken into account at the analysis of geographical variability of vocal behaviour.
2. Material and Methods
Methods of geographical dialects can be applied to songs of birds. For example, sonograms can be used for definition of vocal variants, which are geographically distributed. Frequently (usually) two qualitative methods define (Mundinger, 1982):
• revealing of phonetic distinctions (phoneme) (frequency of a sound - KHz, its form on sonogram – (a pronunciation) – syllabic variability – isophones;
• revealing of lexical distinctions (lexicon) (dictionary structure – change of phrases of songs) – a regional lexicon – isolexes.
However, it is difficult to differentiate (to distinguish) new lexical variants (types of songs, their parts) from extreme (changed) phonetic variants ("figures" of a sound on sonograms) of known syllabic types (that are changed forms of song elements, making a variational line (row) of syllables or elements of one type). As it is difficult to define (determine) a limit at transition of variational lines (rows) of one song type of birds in a variational line (row) of other dialect (modified) song type, which "is obviously close, related" to the first on the origin - so-called "related" vocal lines (rows) at the given species of birds - probably, developed on the basis of the general (common), uniform, initial "ancient" vocal form.
In this work we show the qualitative analysis of song types on above stated methods and we shall try to reveal some features of cultural evolution of chaffinch song (Fringilla coelebs L.) in populations of the European Russia.
Tape records of male song have been made in northwest (Curonian spit, Baltic Sea) (N=153) and in the central part of the European Russia (Zvenigorod, Moscow, Michurinsk) (N=65), distance between which approximately 1000 km. Sonograms of song types were analyzed with the help of computer program Avisoft SASLab Light. In total about five thousand songs are analyzed. Song types were designated (marked) by Latin letters. The songs of one type, which has been recorded in different points (places) of territory were designated (marked) by numbers in ascending order (A1, A2, A3, etc.)
3. Results and Discussion
According to methods of the qualitative analysis of song types - had been signed (allocated) by us two basic kinds of their distinctions: lexical and phonetic.
Lexical variability of chaffinch song types (Fringilla coelebs L.) – distinction of dictionary structure, change of phrases of song types. Proceeding (basing) from samples, by us it has been marked by four ways of its formation (are submitted below).
3.1. Replacement of a Phrase of One Kind by Another in Songs of One Type at Singing
In variants of song types A (fig. 1.1) and I (fig. 1.2), recoded by tape in different points of territory of Curonian spit (biological research station "Fringilla") - the kind of final stroke, characteristic for these types, is replaced with other, usually making song type C (fig. 1.3).
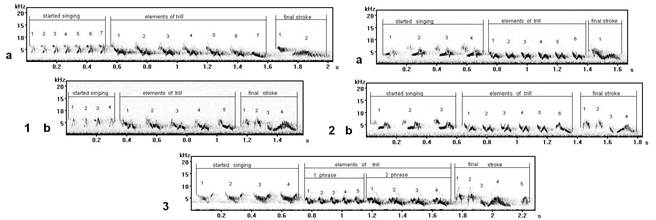
Fig. 1. The songs of one type distinguished (differ) by final strokes (record on Curonian spit, the Kaliningrad region): 1a - song type А8, 1b - song type А5; 2a - song type I 2, 2b - song type I 5; 3 - song type C.
By one type we considered the songs, having two or all three similar parts: started singing (the row of whistle elements), trill elements, final stroke (which they as can be subdivided into phrases – the elements similar under the form).
3.2. Addition of a Phrase to Songs of One Type at Singing
In song type Е1 (fig. 2.1a) a phrase of final stroke is added, and in type М16 (fig. 2.2b) a phrase is duplicated in started singing. Trills in variants of song type M differ because at singing there is an accent on different components of elements. Two such forms of a trill in song type M meet by equal frequency in territory of Curonian spit and characterize individual variability of songs of one type.
3.3. Removal of a Phrase from Songs of One Type at Singing
Song type К2 (fig. 3.1a) it is possible to consider as the uncompleted form of song type I (fig. 3.1b), and song type Q (fig. 3.2a) – such as uncompleted song type D (fig. 3.2b) (on sonograms there is no final stroke). In "incomplete" song types last phrases on hearing are perceived as final stroke (in "full" forms they sound as trill) – therefore in separate, but rare types of songs have been allocated (signed).
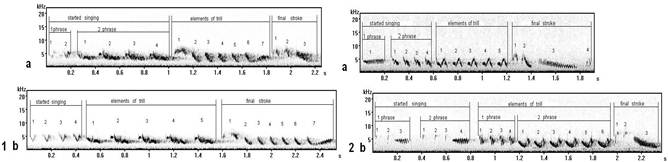
Fig. 2. The songs of one type differing by an additional phrase (record on Curonian spit): 1a - song type Е1, 1b - song type Е2; 2a - song type М12, 2b - song type М16.
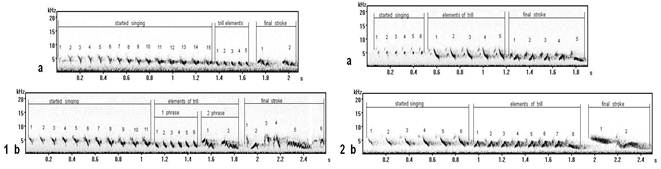
Fig. 3. The samples of songs - similar on the first phrases, but they relate to different types (are recorded on Curonian spit in repertoires of different males): 1a - song type К2, 1b - song type В3; 2a - song type Q, 2b - song type D2.
3.4. Phrase Combinations of Different Song Types at Singing
In repertoires of the some males there were the songs consisting of phrases of other types of songs (fig. 4). Similar cases are described from experiments song training when to caught male of chaffinch in sensitive period daily played different songs of two wild males. In the result, after a while the caught bird sang a song uniting phrases of songs of these two wild males (Nottebohm, 1967; Jellis, 1977). Probably, and by us the marked cases are the certificate of non-standard learning of song types during their crystallization (stabilization).
Such phenomena of lexical distinction of songs of one type within the limits of the certain population of some species sparrow many bioacoustics (scientists) explained from the point of view of natural variations in repertoire of the birds based on vocal generalization and individualization. The phenomena giving development of songs (developmental interaction) can include cooperating processes of imitation (copying) and improvisation (temporarily-dependent copying, a dispersion of young, drift, a regrouping of syllables, phrases) (Lemon, 1975; Burt, Beecher, 2000; Slater et al., 1984).
By song birds are trained from old birds in the first year of a life (Thorpe, 1958), and training (song learning) occurs in most cases more, than from one individual. Various types of songs occur (origin) as a result: migrations, mistakes at training (copying) and improvisation. New song types can be considered as a new growth in a course of "cultural mutation". Distribution of song types in a population corresponds to the assumption, that birds copy them from other individuals of species at random, and that less than 15 % of these new growths influence creation of new song types. One individual of chaffinch male can have repertoire from 1-6 types of songs (Slater, Ince, 1979; Slater et al., 1980).
Thus, the examples of lexical distinctions of song types shown (resulted) by us can be based on mistakes of training and processes of improvisation at singing, that it is possible to name "cultural mutation" in limits of population song cultures. Such cases make small percent from all song types of a population.
Phonetic (syllabic) variability of chaffinch song types (Fringilla coelebs L.) – distinctions in a manner (ways, styles) of performances, pronunciations of syllables, of elements of one type – distinction of their forms on sonograms). We considered change of the form (of pronunciation) of elements of one type in different parts of songs: started singing (the row of whistle elements), a trill (an average part) and final stroke.
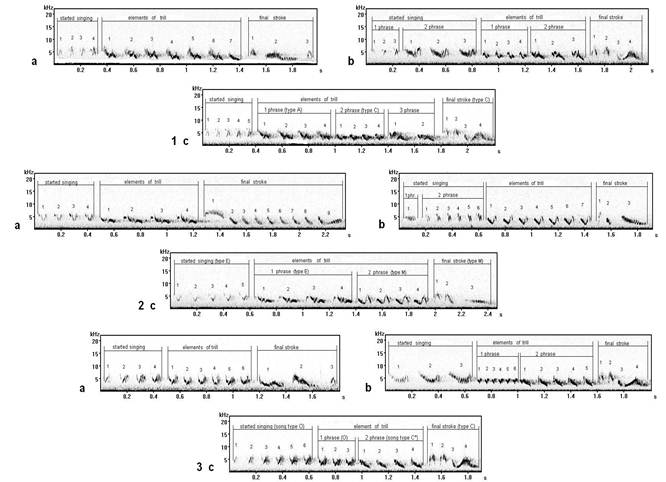
Fig. 4. Sonograms of the combined types of songs at chaffinch (Fringilla coelebs L.), recorded on Curonian spit (those song types - the phrases of which they consist, also are shown): 1a - song type А6, 1b - song type С5, 1c - song type AC; 2a - song type Е6, 2b - song type М1, 2c - song type ЕМ; 3a - song type О3, 3b - song type С*13, 3c - song type P (OC *).
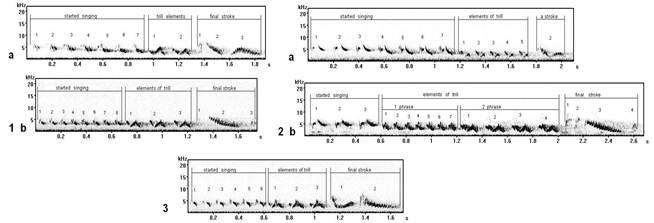
Fig. 5. Sonograms of different types of songs, started singing of which will consist of similar elements under the form, but modified in repertoires at different males (record on Curonian spit): 1a - song type F7, 1b - song type G6; 2a - song type I 9, 2b - song type G1; 3 - song type D1.
3.5. Syllabic Variability in Started Singing of Song Types
Started singing of song types F (fig. 5.1a) and G 6 (fig. 5.1b) are similar in the base (initial) form of elements, but they are sung differently a little. Started singing (the row of whistle elements) in song types I (fig. 5.2a) and G 1 (fig. 5.2b) as are similar, but they also differ in a manner of performance at singing. The song type D (fig. 5.3) has original (distinguished) started singing, but in the general (common) structure of elements « v \ » is similar with started singing of song types (F, G, I) analyzed by us. Thus, even within the limits of a local population (Curonian spit) variability of syllables (elements) of one kind in different types of songs is observed at their different manner of performance (of singing).
It is possible to result (to show) one more example of vocal variability of started singing in different song types (fig. 6).
Elements of started singing (the row of whistle elements) in these song types are arc-similar under the form, but can sound differently: song type С* (fig. 6.2), song type C# (fig. 6.5) have a little creaking (gnash) of a sound "vji-vji", that confirms other pronunciation. In song types Т (fig. 6.4) and S (fig. 6.3) the manner of performance of started singing is similar with the previous song types (C*, C#), but a sound is more thin - "fui-fui". At song type C (fig. 6.1) elements of started singing can sound most precisely, by high tone – "fuit-fuit-fuit".
Thus, despite of similarity of elements in the form (on sonogram), and their similar sound at singing, exists their different pronunciation, a manner of performance, that is phonetic distinction. How these phonetic forms of started singing were fixed in song culture of a population – complex (difficult) question (mistakes at training of songs, folding or forming another song cultures in different local populations, and mixing song traditions at migration?).
Casual copying of song types with possible mistakes, should conduct (lead) to change of birds repertoire of populations during some time. The birds, which were born later, had less time for song training that could conduct (lead) to more erroneous copying (Slater et al., 1980). +
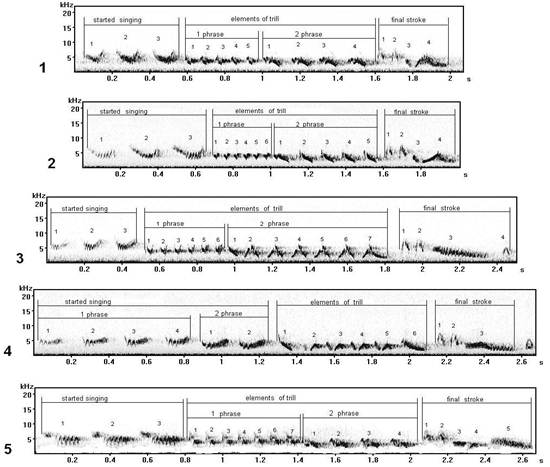
Fig. 6. Sonograms of types of songs, started singing of which will consist of similar elements under the form, but with different manners of their singing (performance): 1 - song type С1 (record in settlement Fishing, Curonian spit), 2 - song type C*, 3 - song type S (record in biostation "Fringilla", Curonian spit), 4 - song type T3 (settlement Fishing, Curonian spit), 5 - song type C* 11 (record in Moscow).
3.6. Syllabic Variability of Elements in Trill of Song Types
Similarity of started singing (the row of whistle elements) in these song types (D, F, G) was already discussed, but if to pay attention on trill elements - it is possible to note their similar base form, but thus to agree - that they differently have been sung in separate song types. In general, the given song types – D (fig. 7.1), F (fig. 7.2), G (fig. 7.3) are similar among themselves, and sometimes their some variants it is difficult to differentiate as three song types, but, nevertheless, their phrases (parts) in many respects differ in a manner of performance of elements at singing. Probably, such phonetic distinctions of the same elements also give different types of songs during cultural evolution of populations (in space and in time).
The trill of these three forms of song type C (fig. 8) consists of two phrases. The first phrase - finer (thin) under the form ("angular") elements, on hearing as a row of thin sounds ("til-til-til"), the second phrase - the complex (difficult) elements consisting of two subelements, and on hearing as more powerful sound ("tel-tel"). On sonograms the given elements both of the first, and of the second phrase have variability, though the initial (base) form of them is uniform, but in different song types at singing have been sung by different styles. The different phonetics (pronunciation) of a trill can generate (give) dialects of songs of one type in different territories, forming vocal culture of a local population.
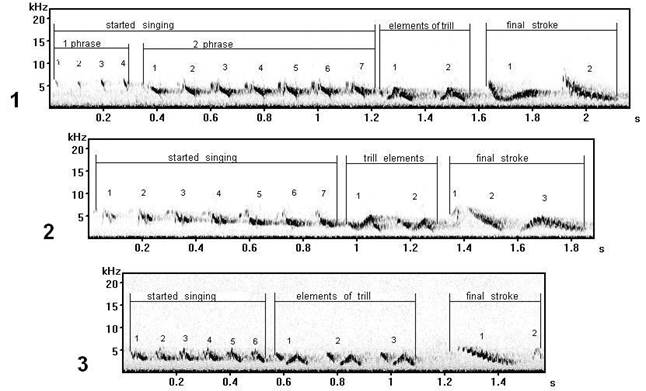
Fig. 7. The types of songs having similar trill elements, but with different ways of their singing execution (performance) at singing (are recorded on Curonian spit): 1 - song type D3; 2 - song type F7; 3 - song type G7.
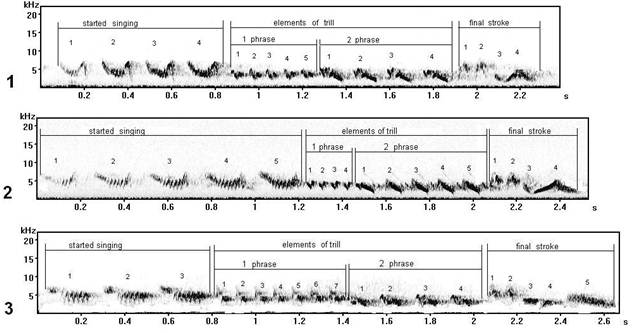
Fig. 8. Vocal variability of trill syllables of song type C: 1 - song type С1 (record in a settlement Fishing, Curonian spit), 2 - song type С*11 (record in settlement Sea, Curonian spit), 3 - song type C* 11 (record in Moscow).
Elements of a trill of song type M (fig. 9) are similar in the base, but differ on a manner of performance at singing: in song types M 1 (fig. 9.1) and M 8 (fig. 9.2) the accent at singing goes on the bottom bend of elements, and in song type M 12 (fig. 9.3) – the accent on the top bend. Such phonetic variability of syllables of a trill of song type M is observed at individuals of chaffinch males (Fringilla coelebs L.) within the limits of one local population (on Curonian spit).
Thus, phonetic variability of syllables (elements) of song types represents their basic (initial) structure, which can be uniform, similar, but modified at singing different chaffinch males (Fringilla coelebs L.) both within the limits of one local population, and in populations in different territories.
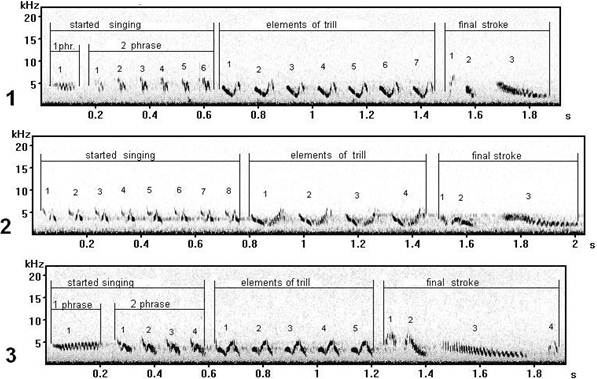
Fig. 9. Vocal variability of trill syllables of song type M (record on Curonian spit): 1 - song type М1, 2 - song type М8, 3 - song type М12.
3.7. Syllabic Variability in Final Stroke of Chaffinch Song Types (Fringilla coelebs L.)
The extreme phonetic variant of song type M (fig. 10.1) is similar to song type J (fig. 10.2). Final strokes are similar on last elements.
We have attributed (related) these songs to one type (fig. 11). But despite of similar general (common) structure, on sonograms precise syllabic variability comes to light, is especial in final stroke: at song type C * (fig. 11.2) last element (which by the "triangular" form at song type C (fig. 11.1)) is divided in three separate subelements. This interesting phonetic difference which changes a kind of a phrase as a whole so, changing also the lexical party (side) of the given song type.
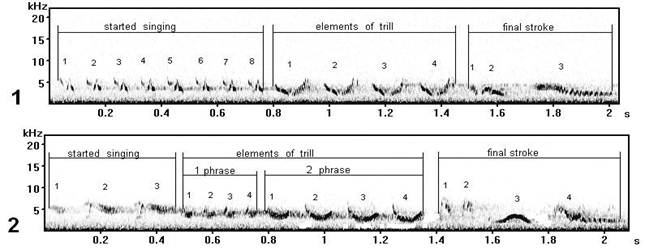
Fig. 10. Vocal variability of final strokes of song types М8 and J6 (record on Curonian spit): 1 - song type М8, 2 - song type J6.
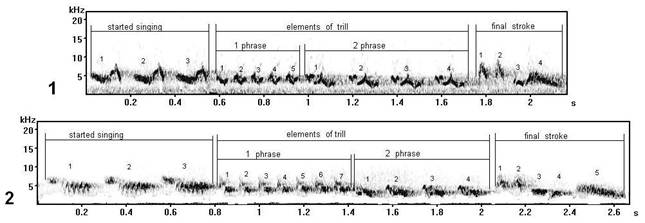
Fig. 11. Vocal variability of final strokes of song type C (record in Moscow): 1 - song type С7, 2 - song type C* 11.
On sonograms a final stroke (fig. 12) has a number of phonetic features: at song type V1 (fig. 12.1) before last element (7) - three small elements of higher frequency (4, 5, 6); the second small elements (2) in final strokes also differ under the form. These phonetic features influences lexicon of song type as a whole, because there is a change of a kind of a phrase. The given samples of song type V have been recorded in repertoires of more than 1-2 individuals of chaffinch (Fringilla coelebs L.) in their local populations.
The case is interesting, when in one population (a settlement Wood or Lesnoy, Curonian spit) songs of one type, but differently performance at singing, have been found at singing of two males (are recoded in the certain day with identical conditions). One of them sang this song type normally (as the majority in our sample) (fig. 13.1), and another (fig. 13.2) - with appreciable errors in phonetics of elements (in their pronunciation) that is reflected in the form on sonogram. In repertoire of everyone male their song form are repeated many times at record (that is already was characteristic for them).
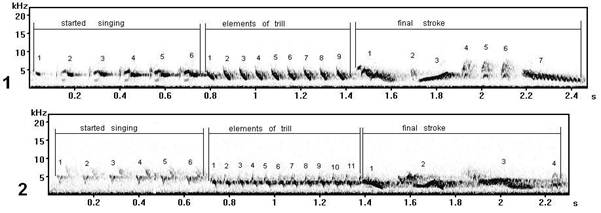
Fig. 12. Vocal variability of final strokes of song type V, recorded in different areas of Russia: 1 - song type V1 (record on Curonian spit, the Kaliningrad region), 2 - song type V (record in Zvenigorod, the Moscow region).
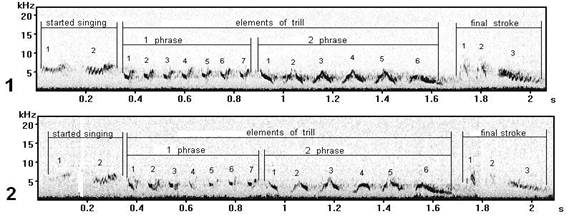
Fig. 13. Normal (1) and (2) forms of song type S taught (learnt) with phonetic mistakes (record in settlement Wood, Curonian spit).
Whether incorrectly learnt form of song type S (fig. 13) will be kept by the subsequent new generation by song training or remain as individual "mutation" which at all will disappear? Whether the erroneous song form becomes sometime of the one of characteristics of song cultures, traditions for one of local populations, probably, already dialect (changed) for that form of this type, which once was norm? It is difficult to answer, it is possible to assume only.
4. Conclusion
Thus, as a result of lexical (concerning phrases or parts of songs) and phonetic (concerning to syllables, elements of song types) vocal variability in local populations of chaffinch (Fringilla coelebs L.) the certain song cultures are formed, which are capable to change during time and can make dialect forms on all area of distribution of a species. For many birds species the patterns of variability represent change of vocal traditions are result of cultural evolution (Mundinger, 1980). Frequently, extreme (changed) phonetic variants turn into lexical variants (phrases vary because of change of forms of elements). In these cases it is difficult to differentiate new lexical variants from extreme phonetic variants of known syllabic types.
Equally with a substantiation of distinctions and variability of song types of birds as product (result) of mistakes of copying, of improvisation at singing and of transfers of features of song traditions to the following generation during cultural evolution (Slater et al., 1979, 1984; Ince et al., 1980), there is an assumption of gradual phylogenetic complication of initial "ancient" (more simple on the structure) song types to more "perfect", complex (difficult) in structure song forms, existing in a population equally with first, on the basis of occurrence of their geographical variability (biomorphism) (Simkin, 1983).
Nevertheless, there are a number of generalizations (Mundinger, 1982; Kroodsma, Miller, 1982):
1. differentiation (division or sharing, distinctions on the basis of the general (common)) of syllables of song types is widely distributed in the nature (Whitney, Miller, 1987; Petronovich, Baptista, 1984; Thielcke, 1984+);
2. microgeographical researches are perspective in phonetic variability, macrogeographical variability includes regional distinctions and similarities in syllabic sets (in groups of types of elements) (that is lexical variability - change of separate song parts and of species specific patterns of song as a whole);
3. among sparrow birds spatial distribution of vocal regional variability can give some forms, the majority of them are general (common), in which various regions were characterized by qualitative distinctions of syllables of songs (different types of song patterns of a species) (Wiley, 1971; Kroodsma, 1974; Nottebohm, 1969; Bertram, 1970; Lemon, 1965, 1966; Avery, Oring, 1977; Grimes, 1974);
4. microgeographical studying of vocal variability focus attention to processes of cultural evolution, macrogeographical studying gives the result in biological evolution;
5. very specific variations (changes) in syntax (sequence, combination of syllables) have been found at some species of birds (Thielcke, Linsenmair, 1963; Kroodsma, 1980; Mundinger, 1975; Bitterbaum, Baptista, 1979);
6. «islands» distribution correlates (have connection) with increase in variability of song patterns; there were the birds - in regular intervals distributed, and during nesting had constantly high density, distinction of their songs – small (insignificant) (Thielcke, 1969).
Mechanisms of forming and development of song traditions of many species of birds remain interesting for clearing (precise) evolutionary directions of species specific song during time and space. Thus it is necessary to note, that at some species the vocalization are not related by social training (song learning), and it is completely hereditary, genetically predetermined. Nevertheless, and these hereditary vocalizations are capable to develop during evolution, can change in the structure. Therefore, probably, influence of heredity and of environment cannot be strengthened in their separate values (accents) – would be more correct to consider forming song of birds at their dynamical interaction.
References
- Avery M., Oring L. W.1977. Song dialect in the Bodolink (Dolichonyx oryzivorus) // Condor. № 79. P. 113-118.
- Bertram B.1970. The vocal behavior of the Indian Hill Mynah (Gracula religiosa) // Anim. Behav. Monogr. № 3. P. 81-192.
- Bitterbaum E., Baptista L. F.1979. Geographical variation in songs of California House Finches (Carpodacus mexicanus) // Auk. № 96. P. 462-474.
- Burt J. M., Beecher M. D.2000. Social influences during song development in the song sparrow: a laboratory experiment simulating field conditions // Animal Behaviour. № 59. P. 1187-1197.
- Grimes L.G.1974. Dialects and geographical variation in the song of the splendid sunbird (Nectarinia coccinigaster) // Ibis. № 116. P. 314-329.
- Ince S. A., Slater P. J. B., Weismann C.1980. Changes with time in the song of a populations Chaffinches // Condor. № 82. P. 285-290.
- Kreutzer M.1974. Stereotypie et varianttions dans les chants de proclamation territoriale chez le Troglodyte (Troglodytes troglodytes) // Rev. Comp.Anim. № 8. P. 270-286.
- Kroodsma D. E.1974. Song learning, dialects, and dispersal in the Bewick`s Wren // Tierpsychol. № 35. P. 352-380.
- Kroodsma D. E. 1980. Winter Wren singing behaviors: A pinnacle of song complexity // Condor. № 82. P. 180-188.
- Kroodsma D. E., Miller E. H., Quellet H.1982.Communication and behavior an interdisciplinary series. Vol. 2: Song leaning and its consequence / D.E. Kroodsma, E.H. Miller. London: Academic press, 347 p.
- Kurath H.1972. Studies in Areal Linguistics. Bloomington: Indiana Univ. Press, 127 p.
- Lemon E. L.1965. The song repertoire of Cardinals (Richmondena cardinals) at London, Ontario // Can. J. Zool. № 43. P. 559-569.
- Lemon E. L.1966. Geographic variation in the song of Cardinals // Can. J. Zool. № 44. P. 413-421.
- Lemon L. E. 1975. How birds develop song dialects // Condor. № 77. P. 385-406. +
- Mundinger P. C.1975. Song dialects and colonization in the House Finch, Carpodacus mexicanus, on the east coast // Condor. № 77. P. 407-422.
- Mundinger P. C.1979. Call learning in the Carduelinae: Ethological and systematic considerations // Syst. Zool. № 28. P. 270-283.
- Mundinger P. C.1980. Animal cultures and a general theory of cultural evolution // Ethol. Sociobiol. № 1. P. 183-223.
- Mundinger P. C.1982. Microgeographic and macrogeographic variation in acquired vocalizations of birds // Acoustic communication in birds. New York, P. 147-208.
- Nottebohm F.1967. The role of sensory feedback in development of avian vocalizations // Proc. Int. Ornithol. Cong. 14. Oxford-Еdinburg. P. 265-280.
- Nottebohm F.1969. The song of the Chingolo, Zonotrichia capensis, in Argentina: Description and evaluation of a system of dialects // Condor. № 71. P. 299-315.
- Petrinovich L., Baptista L. F.1984. Song dialects, mate selection, and breeding success in white-crowned sparrows // Anim. Behav. № 32. P. 1078-1088. +
- Simkin G. N.1983. Typological organization and population phylogeny of song at birds // Moscow group of nature science (Department of Biology, Vol. 88, № 1). P. 15-27. +
- Slater P. J. B., Ince S. A.1979. Cultural evolution in chaffinch song // Behaviour. № 71. P. 146-166.
- Slater P. J., Ince S. A., Colgan P. W.1980. Chaffinch song types: their frequencies in the population and distribution between repertoires of different individuals // Behaviour. № 75. P. 207-218.
- Slater P. J. B., Clement F. A., Goodfellow D. J.1984. Local and regional variations in chaffinch song and the question of dialects // Behaviour. № 88. P. 76-97. +
- Thielcke G., Linsenmair K. E.1963.Zur GeographiachenVariation des Gesanges des Zilpzalpa, Phylloscopus collybita, in Mittel und Südwesteuropa mit einem Vergleich des Gesanges der Fitis, Phylloscopus trochilus // Journ. ornithol.№ 104. P. 372-402.
- Thielcke G.1965. Gesangsgeographische Variation des Gartenbaumläufers (Certhia brachydactyla) im Hinblick auf das Artbildungsproblem // Z. Tierpsychol. № 22. P. 542-566.
- Thielcke G.1984. Gesangslernrn beim Gartenbaumläufer Certhia brachydactyla // Vogelwarte. № 32. P. 282-297.
- Wiley R. H.1971. Song groups in a singing assembly of Little Hermits // Condor. № 73 P. 28-35.
- Whitney C. L., Miller J.1987. Song leaning in the wood thrush // Can. J. Zool. № 65. P. 1038-1042. ++



